S-ar putea să vă placă și
- Sistema Retículo EndotelialDocument5 paginiSistema Retículo EndotelialJoshuaNeftalí100% (1)
- La Rutina de Entrenamiento para Perder PesoDocument11 paginiLa Rutina de Entrenamiento para Perder PesoKarina ManzanelliÎncă nu există evaluări
- Ciclo CardiacoDocument2 paginiCiclo CardiacoDelia Katherine Tuesta QuispeÎncă nu există evaluări
- Tiempo de SangríaDocument2 paginiTiempo de Sangríapayaso7Încă nu există evaluări
- Resumen Abdomen Anato AtlasDocument14 paginiResumen Abdomen Anato AtlasPedro Custodio SánchezÎncă nu există evaluări
- Tejido Excito ConductorDocument21 paginiTejido Excito ConductorVictoria Bazurto100% (1)
- Robin de Texas - Ases Del Oeste 327 Ed B - Keith LugerDocument157 paginiRobin de Texas - Ases Del Oeste 327 Ed B - Keith LugerJC NiNuÎncă nu există evaluări
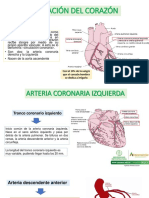
- CorazónDocument12 paginiCorazónAlexander Hernandez100% (1)
- Inmunidad AdquiridaDocument21 paginiInmunidad AdquiridaRoberto Rosas100% (1)
- Gasto CardíacoDocument8 paginiGasto Cardíacopanch0man100% (9)
- Patrones Radiológicos PulmonaresDocument7 paginiPatrones Radiológicos PulmonaresJohn Maccapa ChancaÎncă nu există evaluări
- Histología Del CorazonDocument48 paginiHistología Del CorazonVicToRia86% (7)
- Fisiología de Guyton Resumen Capítulo 5Document2 paginiFisiología de Guyton Resumen Capítulo 5Pablo Andrés Zurita75% (20)
- Capitulo 9 Resumen Guyton e Hall Tratado de Fisiologia MedicaDocument14 paginiCapitulo 9 Resumen Guyton e Hall Tratado de Fisiologia MedicaGilberto F. Fernandez100% (2)
- Resumen Guyton Capitulo 28Document5 paginiResumen Guyton Capitulo 28Carol Jennifer Núñez Campos100% (1)
- Anatomía Del PulmónDocument28 paginiAnatomía Del PulmónDaniela Gómez Sifuentes50% (2)
- Fisiología de Guyton Resumen Capítulo 3Document4 paginiFisiología de Guyton Resumen Capítulo 3Pablo Andrés Zurita88% (8)
- Capitulo 17 Guyton ResumenDocument16 paginiCapitulo 17 Guyton ResumenrubenÎncă nu există evaluări
- Resumen de Electrocardiograma Normal Capitulo 11 de Fisiologia de Guyton 72986 Downloable 744015Document6 paginiResumen de Electrocardiograma Normal Capitulo 11 de Fisiologia de Guyton 72986 Downloable 744015nathalia cabrera100% (1)
- Capitulo 40 GuytonDocument3 paginiCapitulo 40 GuytonGaby GarridoÎncă nu există evaluări
- Anatomia de Los PulmonesDocument24 paginiAnatomia de Los PulmonesKatleen Rodas0% (1)
- Resumen Capitulo 35 FisiologiaDocument6 paginiResumen Capitulo 35 FisiologiaSofíaMelo100% (2)
- GUYTON-CAPÍTULO 10 - Excitación Rítmica Del CorazónDocument3 paginiGUYTON-CAPÍTULO 10 - Excitación Rítmica Del CorazónKAREN ANDREA GARCIA RAMOSÎncă nu există evaluări
- Capitulo 14 de Guyton ResumenDocument7 paginiCapitulo 14 de Guyton Resumenor agÎncă nu există evaluări
- Fisiologia Del Sistema Cardiovascular Parte IDocument32 paginiFisiologia Del Sistema Cardiovascular Parte IJennifer Dayana Roa Umaña100% (1)
- Grandes VasosDocument51 paginiGrandes VasosEmmylly Barbosa100% (1)
- Flujo Sanguineo Muscular y Gasto CardiacoDocument37 paginiFlujo Sanguineo Muscular y Gasto CardiacoMeztliMagañaÎncă nu există evaluări
- Guyton-Capítulo 9-Músculo CardíacoDocument6 paginiGuyton-Capítulo 9-Músculo CardíacoKaren García RamosÎncă nu există evaluări
- Examen Físico CardiovascularDocument27 paginiExamen Físico CardiovascularCarmenÎncă nu există evaluări
- Fisiología de Guyton Resumen Capítulo 8Document2 paginiFisiología de Guyton Resumen Capítulo 8Pablo Andrés Zurita86% (21)
- Resumen Cap 39Document7 paginiResumen Cap 39jonasgs_92100% (2)
- Capitulo 21. Flujo Sanguineo Muscular y Gasto Cardiaco Durante El Ejercicio, La Circulacion Coronaria y La Cardiopatia IsquemicaDocument32 paginiCapitulo 21. Flujo Sanguineo Muscular y Gasto Cardiaco Durante El Ejercicio, La Circulacion Coronaria y La Cardiopatia IsquemicaFaBian AŋDrIk100% (2)
- Anastomosis Porto-CavaDocument1 paginăAnastomosis Porto-CavaFlor Veiga100% (2)
- Irrigación Del CorazónDocument12 paginiIrrigación Del CorazónVALERIE KUHLMANN CHABUTÎncă nu există evaluări
- Visión General de La CirculaciónDocument8 paginiVisión General de La CirculaciónHéctor BacÎncă nu există evaluări
- Fisiologia Cardiaca GuytonDocument20 paginiFisiologia Cardiaca Guytonmate la femina100% (1)
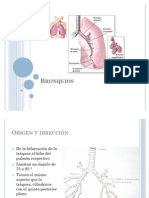
- Anatomía de BronquiosDocument8 paginiAnatomía de Bronquiosprincezithalinda0150% (2)
- Resumen Capitulo V Guyton.Document8 paginiResumen Capitulo V Guyton.Ivette Diaz100% (1)
- Mecanica de Materiales FinalDocument48 paginiMecanica de Materiales FinalDalliana Malu Soplapuco AguilarÎncă nu există evaluări
- Resumen de Capitulo 14 de Fisiologia de Guyton 120 EdDocument2 paginiResumen de Capitulo 14 de Fisiologia de Guyton 120 EdLuis Víctor Bamtz0% (1)
- Cap. 27 Filtración GlomerularDocument4 paginiCap. 27 Filtración GlomerularJocelyne Benito100% (2)
- Jeet Kune Do - Bruce LeeDocument46 paginiJeet Kune Do - Bruce LeeDavidPinedaAlvaradoÎncă nu există evaluări
- Excitación Rítmica Del CorazónDocument8 paginiExcitación Rítmica Del Corazónfalco8molinaÎncă nu există evaluări
- Excitacion Ritmica Del CorazonDocument20 paginiExcitacion Ritmica Del CorazongloriaÎncă nu există evaluări
- Cap. 10. Excitación y Conducción Del CorazónDocument32 paginiCap. 10. Excitación y Conducción Del CorazónPaulo Cesar Silva RibeiroÎncă nu există evaluări
- Capitulo 12 Guyton EkgDocument7 paginiCapitulo 12 Guyton EkgDanielaNuñezGonzalez100% (2)
- Capitulo 7 Guyton ResumenDocument4 paginiCapitulo 7 Guyton ResumenEliezer Sop75% (4)
- Analisisdel Capitulo 15 Fisiologia GuytonDocument8 paginiAnalisisdel Capitulo 15 Fisiologia GuytonBryan LópezÎncă nu există evaluări
- Capitulo 11: Electrocardiograma Normal - GuytonDocument3 paginiCapitulo 11: Electrocardiograma Normal - GuytonAlexis Andrade Ibarra100% (3)
- Guyton Capitulo 11 DiapositivaDocument5 paginiGuyton Capitulo 11 DiapositivaFrancisco Gabriel Gervacio0% (1)
- Excitacion Ritmica Del CorazonDocument24 paginiExcitacion Ritmica Del CorazonAngel LopezÎncă nu există evaluări
- Arterias Elásticas y Venas de Gran CalibreDocument5 paginiArterias Elásticas y Venas de Gran CalibreIsrael SarmientoÎncă nu există evaluări
- Vascularización e Inervación Del CorazónDocument3 paginiVascularización e Inervación Del CorazónPierina Salguero100% (1)
- Relaciones de Los Pulmones y de La PleuraDocument38 paginiRelaciones de Los Pulmones y de La PleuraGustavo Rosario Rojas0% (2)
- Vascularizacion e Inervacion de Abdomen PDFDocument24 paginiVascularizacion e Inervacion de Abdomen PDFbellaaaaaaÎncă nu există evaluări
- Inervacion e Irrigacion de Los Pulmones y ParenquimaDocument31 paginiInervacion e Irrigacion de Los Pulmones y ParenquimaDaniel Nieves100% (1)
- Resumen Murray Microbiologia Capitulo 10Document6 paginiResumen Murray Microbiologia Capitulo 10Paula Nicole Torres Retamal100% (1)
- Fisiología-Excitación Rítmica Del CorazónDocument19 paginiFisiología-Excitación Rítmica Del CorazónDiana América Chávez Cabrera100% (1)
- Irrigación CardiacaDocument3 paginiIrrigación CardiacaJhonatan Villa100% (1)
- Hipofisis HistologiaDocument6 paginiHipofisis HistologiaAníbal CIÎncă nu există evaluări
- Ciclo CardiacoDocument2 paginiCiclo CardiacoValentina FuquenÎncă nu există evaluări
- Capítulo 9: Músculo Cardíaco: El Corazón Como Bomba y La Función de Las Válvulas CardíacasDocument37 paginiCapítulo 9: Músculo Cardíaco: El Corazón Como Bomba y La Función de Las Válvulas CardíacasManuela Monsalve TamarizÎncă nu există evaluări
- UABP 2. Músculo Cardíaco y Ciclo CardíacoDocument22 paginiUABP 2. Músculo Cardíaco y Ciclo CardíacoMartín PanicoÎncă nu există evaluări
- Musculo Cardiaco ClaseDocument19 paginiMusculo Cardiaco ClaseOscarChávezSánchez100% (2)
- Máxima Goleada Del Clásico PlatenseDocument6 paginiMáxima Goleada Del Clásico PlatenseScorpion1985Încă nu există evaluări
- Tema VII El MusculoDocument42 paginiTema VII El MusculoandoporÎncă nu există evaluări
- Rev79 Art4Document25 paginiRev79 Art4Facundo AlmadaÎncă nu există evaluări
- Musculos Expresion FacialDocument4 paginiMusculos Expresion FacialJohana Elizabeth VelascoÎncă nu există evaluări
- Rescate Vertical, Cuerdas y NudosDocument29 paginiRescate Vertical, Cuerdas y NudosPasion Bomberil100% (2)
- Espanol para Nosotros Chapter 7Document34 paginiEspanol para Nosotros Chapter 7Earth003Încă nu există evaluări
- Armas de Artes MarcialesDocument7 paginiArmas de Artes MarcialesJuan Jacobo Janeiro OnorezÎncă nu există evaluări
- Guia Voleibo 4to Periodo Grado 7Document3 paginiGuia Voleibo 4to Periodo Grado 7Marta Beatriz Verbel ArroyoÎncă nu există evaluări
- Formato UNTREF para Trabajos de Investigación Científicos 2010Document4 paginiFormato UNTREF para Trabajos de Investigación Científicos 2010Andrés LarrahonaÎncă nu există evaluări
- Danza - Ciclo III - Tema 2 (Terminología)Document2 paginiDanza - Ciclo III - Tema 2 (Terminología)César Arturo Aguilar BocanegraÎncă nu există evaluări
- Tema 2 - Problemas Cinemática - 1213Document20 paginiTema 2 - Problemas Cinemática - 1213Juan Dario Ruiz ZapataÎncă nu există evaluări
- Larson Paginas y Ejercicios Seleccionados Hasta Prim ParcialDocument1 paginăLarson Paginas y Ejercicios Seleccionados Hasta Prim ParcialMantenimiento MECÎncă nu există evaluări
- TrabajoDocument114 paginiTrabajoJohn Jairo Garcìa CèspedesÎncă nu există evaluări
- Pruebas Tacticas de Campo NoDocument3 paginiPruebas Tacticas de Campo NoJaime A. VilladaÎncă nu există evaluări
- Compendio Del VoleibolDocument17 paginiCompendio Del VoleiboltrogloditaÎncă nu există evaluări
- Centros AcuáticosDocument16 paginiCentros AcuáticosDenise RetloveÎncă nu există evaluări
- Estadios en México y El MundoDocument28 paginiEstadios en México y El MundoOswaldo MondragonÎncă nu există evaluări
- Diagnóstico y Propuestas de Accesibilidad Universal y Diseño para Todos en El Parque de Los Niños de BucaramangaDocument17 paginiDiagnóstico y Propuestas de Accesibilidad Universal y Diseño para Todos en El Parque de Los Niños de BucaramangaLeonardo Enrique Díaz SuárezÎncă nu există evaluări
- Informe de FisicaDocument9 paginiInforme de FisicaLeticia OrellanaÎncă nu există evaluări
- Pa5 HotelDocument14 paginiPa5 HotelLisbete ZegarraÎncă nu există evaluări
- Folleto-Summer-Camp 2024Document10 paginiFolleto-Summer-Camp 2024Kennedy JadelÎncă nu există evaluări
- Parrilla Del 18/09/10Document2 paginiParrilla Del 18/09/10Xornal de GaliciaÎncă nu există evaluări
- TENISDocument3 paginiTENISyoyoÎncă nu există evaluări
- Span 1 Sem Test Review CH 8 and 9Document2 paginiSpan 1 Sem Test Review CH 8 and 9lasalernoÎncă nu există evaluări
- AUTORIZACIÓNDocument2 paginiAUTORIZACIÓNerika mamani huamanÎncă nu există evaluări
- Ver en MapaDocument7 paginiVer en MapaAdalin OlivaresÎncă nu există evaluări