S-ar putea să vă placă și
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De la EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Încă nu există evaluări
- Mecanismos de TransporteDocument4 paginiMecanismos de TransporteJonathan MartínezÎncă nu există evaluări
- Membrana plasmática: estructura, funciones y transporteDocument62 paginiMembrana plasmática: estructura, funciones y transporteAlejandra MuñozÎncă nu există evaluări
- TRANSPORTE A Través de La MEMBRANADocument24 paginiTRANSPORTE A Través de La MEMBRANALaura MoyaÎncă nu există evaluări
- Biofisica, Profesorado de BiologíaDocument21 paginiBiofisica, Profesorado de Biologíacristian silvaÎncă nu există evaluări
- Transporte de Sustancias A Través de Las Membranas CelularesDocument6 paginiTransporte de Sustancias A Través de Las Membranas CelularesAdrianaÎncă nu există evaluări
- Mecanismos de Transporte de Membrana CelularDocument15 paginiMecanismos de Transporte de Membrana CelularAlejandraSánchezÎncă nu există evaluări
- Tipos de Transporte CelularDocument28 paginiTipos de Transporte CelularMisuzu Kamio50% (2)
- Tema 4.Document7 paginiTema 4.Claudia Vazquez GarciaÎncă nu există evaluări
- Transporte Activo PerlaDocument19 paginiTransporte Activo PerlaPerla HernandezÎncă nu există evaluări
- Transporte - BiofisicaDocument15 paginiTransporte - BiofisicaAsucena MarcosÎncă nu există evaluări
- Guión Expo. Transportes MCDocument16 paginiGuión Expo. Transportes MCEtyamadnessÎncă nu există evaluări
- Tema 4Document6 paginiTema 4miguel dizÎncă nu există evaluări
- Cinética vegetalDocument4 paginiCinética vegetalCristhine Pilco Calizaya67% (3)
- Transporte membranas celularesDocument31 paginiTransporte membranas celularesCamila Vargas VeraÎncă nu există evaluări
- Mecanismos de Transporte TransmembranalDocument12 paginiMecanismos de Transporte TransmembranalBerenice Infante100% (1)
- Transporte a través de la membrana celularDocument8 paginiTransporte a través de la membrana celularMariana ReyesÎncă nu există evaluări
- Resumen de Biologia, Principio de Transporte de MembranaDocument3 paginiResumen de Biologia, Principio de Transporte de MembranaKamila AlbaÎncă nu există evaluări
- Transporte de SustanciasDocument4 paginiTransporte de SustanciasKeyla AlmeidaÎncă nu există evaluări
- Tema 2Document8 paginiTema 2juanmideluxeÎncă nu există evaluări
- Transporte Activo y PasivoDocument4 paginiTransporte Activo y PasivoMarley CajamarcaÎncă nu există evaluări
- Biologia Transorte ActivoDocument4 paginiBiologia Transorte ActivojosehlatorreÎncă nu există evaluări
- Transporte A Través de La Membrana PlasmáticaDocument17 paginiTransporte A Través de La Membrana PlasmáticaCarola DJÎncă nu există evaluări
- TIPOS de TRANSPORTE de MembranaDocument14 paginiTIPOS de TRANSPORTE de MembranaAbigail MLÎncă nu există evaluări
- Resumen FisioDocument50 paginiResumen FisioDebb CespedesÎncă nu există evaluări
- Fisiología 1Document19 paginiFisiología 1Rocío TempranoÎncă nu există evaluări
- Membrana y Mecanismo de TransporteDocument3 paginiMembrana y Mecanismo de TransporteCandela PaccotÎncă nu există evaluări
- Mecanismos de Transporte Activo y PasivoDocument4 paginiMecanismos de Transporte Activo y PasivoJacob Diaz100% (3)
- Apuntes de Nefrología Crítica PDFDocument115 paginiApuntes de Nefrología Crítica PDFMiguel Chung SangÎncă nu există evaluări
- 3 Membrana y TransporteDocument32 pagini3 Membrana y TransporteMaria Jose ROÎncă nu există evaluări
- Mhs Tramo Dos AlafDocument59 paginiMhs Tramo Dos AlafZiaraÎncă nu există evaluări
- Transporte CelularDocument8 paginiTransporte CelularCesar Manuel Morales AbreuÎncă nu există evaluări
- 05-Transporte ApunteDocument11 pagini05-Transporte ApunteRODRIGO ACUÑAÎncă nu există evaluări
- Transporte Activo en Epitelio IntestinalDocument19 paginiTransporte Activo en Epitelio IntestinalAlexandra MusajaÎncă nu există evaluări
- 04 Mecanismos de TransporteDocument40 pagini04 Mecanismos de TransporteDANIELA CONSTANZA PEREZ AGUILARÎncă nu există evaluări
- Guía I Membranas Biologicas y Mecanismos de TransporteDocument8 paginiGuía I Membranas Biologicas y Mecanismos de TransporteConstanza NicoleÎncă nu există evaluări
- Conductos Y Bombas de Membrana: Escuela Profesional de Biologia-Bioquimica IiDocument63 paginiConductos Y Bombas de Membrana: Escuela Profesional de Biologia-Bioquimica IiClinton LunaÎncă nu există evaluări
- MR Transporte Activo y PasivoDocument15 paginiMR Transporte Activo y PasivoNahom'ys YoxmanÎncă nu există evaluări
- S03C04 - Membrana (II)Document34 paginiS03C04 - Membrana (II)Alejandro B. SilvaÎncă nu există evaluări
- TransporteDocument12 paginiTransporteTom I. MapacheÎncă nu există evaluări
- Transportes de membranaDocument10 paginiTransportes de membranaDavid PinanjotÎncă nu există evaluări
- Transporte Act y PasDocument3 paginiTransporte Act y PasMarlene JorgeÎncă nu există evaluări
- Transporte de Sustancias A Través de La Membrana Capitulo 4 GuytonDocument26 paginiTransporte de Sustancias A Través de La Membrana Capitulo 4 GuytonFatima QuintanaÎncă nu există evaluări
- Solemne 1 Fisio GenealDocument108 paginiSolemne 1 Fisio GenealGabriela VelásquezÎncă nu există evaluări
- El Proceso de Transporte Es Importante para La Célula Porque Le Permite Expulsar de Su Interior Los Desechos Del Metabolismo y Adquirir NutrientesDocument4 paginiEl Proceso de Transporte Es Importante para La Célula Porque Le Permite Expulsar de Su Interior Los Desechos Del Metabolismo y Adquirir Nutrientesjuanmanuel26Încă nu există evaluări
- Nivelacion Grado SeptimoDocument7 paginiNivelacion Grado Septimoccnlasalle100% (1)
- Estructura de La Membrana Celular y Sus FuncionesDocument63 paginiEstructura de La Membrana Celular y Sus FuncionesAndreaÎncă nu există evaluări
- Fisiologia CelularDocument26 paginiFisiologia Celularanydeyra25Încă nu există evaluări
- Difusion y TransporteDocument7 paginiDifusion y TransporteSam Silguera EspinalÎncă nu există evaluări
- Transporte TransepitelialDocument21 paginiTransporte Transepitelialmedicina crecimientoÎncă nu există evaluări
- Copia de TEMA 3-Transporte A Través de Membranas CelularesDocument21 paginiCopia de TEMA 3-Transporte A Través de Membranas CelularessilviaahernaandezÎncă nu există evaluări
- Clase A Clase Semana 3.2Document13 paginiClase A Clase Semana 3.2DragmxÎncă nu există evaluări
- Transporte celular-Ósmosis,difusión,activoDocument6 paginiTransporte celular-Ósmosis,difusión,activoFco ZalgadoÎncă nu există evaluări
- Copia de TEMA 3-Transporte A Través de Membranas CelularesDocument21 paginiCopia de TEMA 3-Transporte A Través de Membranas CelularessilviaahernaandezÎncă nu există evaluări
- Potencial de Accion CardiacoDocument20 paginiPotencial de Accion CardiacoJean José Reyes BenítezÎncă nu există evaluări
- Medios de Transporte CelularDocument4 paginiMedios de Transporte CelularKevin AlexisÎncă nu există evaluări
- Procesos de Transporte en Las CélulasDocument24 paginiProcesos de Transporte en Las CélulasJuly Paulin BarraganÎncă nu există evaluări
- Mecanismos de Transporte FisioDocument2 paginiMecanismos de Transporte Fisiosiguasale06Încă nu există evaluări
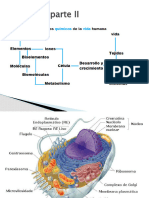
- Copia de La Celula Parte IIDocument71 paginiCopia de La Celula Parte IIp7pm4vsrvnÎncă nu există evaluări
- Transporte de SustanciasDocument77 paginiTransporte de SustanciasClaudia Vargas AndinoÎncă nu există evaluări
- Final EmpresasDocument24 paginiFinal EmpresasDaniel FallaÎncă nu există evaluări
- Diseños de Sistema de RiegoDocument3 paginiDiseños de Sistema de RiegoDaniel FallaÎncă nu există evaluări
- 1405 2768 Polib 45 181 - 5Document15 pagini1405 2768 Polib 45 181 - 5Daniel FallaÎncă nu există evaluări
- El Frijol Caupí,: Recuperación Y Seguridad Alimentaria Pos Emergencias Por LluviasDocument24 paginiEl Frijol Caupí,: Recuperación Y Seguridad Alimentaria Pos Emergencias Por LluviasDaniel FallaÎncă nu există evaluări
- Proyecto Elemento Faltante Maiz 2015Document36 paginiProyecto Elemento Faltante Maiz 2015Daniel FallaÎncă nu există evaluări
- Cultivo in Vitro PDFDocument8 paginiCultivo in Vitro PDFDaniela GiraudinÎncă nu există evaluări
- 1405 2768 Polib 45 181 - 5Document15 pagini1405 2768 Polib 45 181 - 5Daniel FallaÎncă nu există evaluări
- Propagación in VitroDocument33 paginiPropagación in VitroBertha Escobedo RubioÎncă nu există evaluări
- MarschnerDocument733 paginiMarschnerDaniel FallaÎncă nu există evaluări
- Aprendemos Sobre La Utilidad de Las Razones TrigonométricasDocument5 paginiAprendemos Sobre La Utilidad de Las Razones TrigonométricasDaniel FallaÎncă nu există evaluări
- 144938201432610323Document102 pagini144938201432610323bitnerÎncă nu există evaluări
- 4 Pra Cticas Biotecnologi A VegetalDocument45 pagini4 Pra Cticas Biotecnologi A VegetalDaniel FallaÎncă nu există evaluări
- ARROZDocument3 paginiARROZDaniel FallaÎncă nu există evaluări
- Cultivo in Vitro PDFDocument8 paginiCultivo in Vitro PDFDaniela GiraudinÎncă nu există evaluări
- Art 12Document9 paginiArt 12Daniel FallaÎncă nu există evaluări
- Appcacap PDFDocument12 paginiAppcacap PDFDaniel FallaÎncă nu există evaluări
- Manejo integrado plagas enfermedades papaDocument28 paginiManejo integrado plagas enfermedades papaAnonymous 6Jn4hM0Încă nu există evaluări
- Appcacap PDFDocument12 paginiAppcacap PDFDaniel FallaÎncă nu există evaluări
- 032 B PapaDocument35 pagini032 B PapaHéctor Jesús Pacheco PonceÎncă nu există evaluări
- Propagación in VitroDocument33 paginiPropagación in VitroBertha Escobedo RubioÎncă nu există evaluări
- Propagación in VitroDocument33 paginiPropagación in VitroBertha Escobedo RubioÎncă nu există evaluări
- Propagación in VitroDocument33 paginiPropagación in VitroBertha Escobedo RubioÎncă nu există evaluări
- CACAODocument9 paginiCACAODaniel FallaÎncă nu există evaluări
- Manejo integrado plagas enfermedades papaDocument28 paginiManejo integrado plagas enfermedades papaAnonymous 6Jn4hM0Încă nu există evaluări
- Propagación in VitroDocument33 paginiPropagación in VitroBertha Escobedo RubioÎncă nu există evaluări
- CARATULADocument2 paginiCARATULADaniel FallaÎncă nu există evaluări
- 032 B PapaDocument35 pagini032 B PapaHéctor Jesús Pacheco PonceÎncă nu există evaluări
- CACAODocument9 paginiCACAODaniel FallaÎncă nu există evaluări
- Quemado en ArrozDocument3 paginiQuemado en ArrozDaniel FallaÎncă nu există evaluări
- CARATULADocument2 paginiCARATULADaniel FallaÎncă nu există evaluări
- Exposición AgroclimatologíaDocument15 paginiExposición AgroclimatologíaDaniel AlonsoÎncă nu există evaluări
- Drunvalo Melchizedek Serpiente de LuzDocument211 paginiDrunvalo Melchizedek Serpiente de Luzolaolaolaola123100% (1)
- Patogenia de las principales infecciones viralesDocument28 paginiPatogenia de las principales infecciones virales05-EF-HU-ELOYZA URIETH GOMEZ PONCEÎncă nu există evaluări
- Protocolo 14Document6 paginiProtocolo 14Andrea100% (1)
- RespiraciónDocument3 paginiRespiraciónAgus AquinoÎncă nu există evaluări
- RELAJACIÓN DE JACOBSON 16 Grupos MuscularesDocument3 paginiRELAJACIÓN DE JACOBSON 16 Grupos MuscularesMaríaÎncă nu există evaluări
- Importancia Del ColagenoDocument9 paginiImportancia Del ColagenoFrank SanchezÎncă nu există evaluări
- 72 Sanmiguel - Callejon SucreDocument63 pagini72 Sanmiguel - Callejon SucreJuariDocs100% (3)
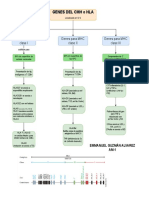
- Genes de HLA o MHCDocument1 paginăGenes de HLA o MHCEmmanuel GÎncă nu există evaluări
- Fenómenos Cadavéricos TransformadoresDocument20 paginiFenómenos Cadavéricos TransformadoresmariÎncă nu există evaluări
- Tesis LGH-Final-1Document89 paginiTesis LGH-Final-1Selene CotardoÎncă nu există evaluări
- Aislamiento de PsicrofilosDocument3 paginiAislamiento de PsicrofilosSara Mucha BeraunÎncă nu există evaluări
- HEMOFILIADocument52 paginiHEMOFILIAMonse PeraltaÎncă nu există evaluări
- S02.s2 Práctica1 DañoCerebralCongénito FlorHuarcaDocument8 paginiS02.s2 Práctica1 DañoCerebralCongénito FlorHuarcaFlor HuarcaÎncă nu există evaluări
- Sanación AngelicalDocument9 paginiSanación AngelicalYamila Lugones83% (6)
- Banco de Preguntas - Esfm - 2018Document93 paginiBanco de Preguntas - Esfm - 2018Franz Ever Quispecahuana Limachi80% (111)
- Hoja de Vida DouglasDocument5 paginiHoja de Vida DouglasEli BatistaÎncă nu există evaluări
- Plaguicidas. Clasificacion QuimicaDocument20 paginiPlaguicidas. Clasificacion QuimicaHéctor Velásquez AlcántaraÎncă nu există evaluări
- Laboratorios acreditados IDEAM matriz aireDocument98 paginiLaboratorios acreditados IDEAM matriz aireJavier Perez NIñoÎncă nu există evaluări
- Programa Biología (Admisión Agronomía UNA Py)Document2 paginiPrograma Biología (Admisión Agronomía UNA Py)Manu RodrigoÎncă nu există evaluări
- Etapas de La Reproducción: Unidad 3Document13 paginiEtapas de La Reproducción: Unidad 3isabel carrilloÎncă nu există evaluări
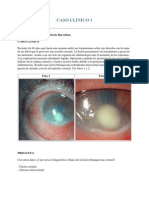
- Caso Clinico OftalmoDocument20 paginiCaso Clinico Oftalmoleyva_777100% (2)
- A Cabeza Descalza - Brian W. AldissDocument159 paginiA Cabeza Descalza - Brian W. AldissJeffrey T. Spaulding100% (1)
- Dermapen PDFDocument10 paginiDermapen PDFisabel rodriguez100% (3)
- VSG-Historia-UtilidadDocument6 paginiVSG-Historia-UtilidadSheila ReyesÎncă nu există evaluări
- Fases de La Marcha HumanaDocument8 paginiFases de La Marcha Humanafederico saldariniÎncă nu există evaluări
- Exposición. Práctica #6. Glutatión y Su Rol OxidativoDocument27 paginiExposición. Práctica #6. Glutatión y Su Rol OxidativoStefani RSÎncă nu există evaluări
- Circulo de Estudio - Area Comunicacion PDFDocument71 paginiCirculo de Estudio - Area Comunicacion PDFBilly Deybi Niño MoralesÎncă nu există evaluări
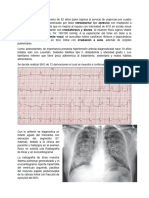
- Caso Clínico de Intregración Hemodinámica y Electrica 2Document17 paginiCaso Clínico de Intregración Hemodinámica y Electrica 2Natalia RodríguezÎncă nu există evaluări
- Principales procesos silvícolas y checklist de maquinariaDocument10 paginiPrincipales procesos silvícolas y checklist de maquinariapedro cÎncă nu există evaluări