S-ar putea să vă placă și
- DD Work - DecDocument3 paginiDD Work - DecFrancis KumarÎncă nu există evaluări
- Audit Report 2016-17Document6 paginiAudit Report 2016-17Francis KumarÎncă nu există evaluări
- 009 Naa Kutumbam 01 25 PDFDocument157 pagini009 Naa Kutumbam 01 25 PDFtelugubad70% (94)
- Requesting For Donation LetterDocument1 paginăRequesting For Donation LetterFrancis KumarÎncă nu există evaluări
- Requesting For Donation LetterDocument1 paginăRequesting For Donation LetterFrancis KumarÎncă nu există evaluări
- Requesting For Donation LetterDocument1 paginăRequesting For Donation LetterFrancis KumarÎncă nu există evaluări
- VISA Foundation 1Document28 paginiVISA Foundation 1Francis KumarÎncă nu există evaluări
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (587)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (265)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (119)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
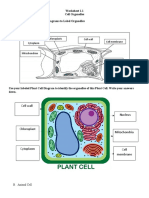
- Worksheet 1.1 Cell OrganelleDocument2 paginiWorksheet 1.1 Cell OrganelleCyndel TindoyÎncă nu există evaluări
- DNA Found in FoodDocument22 paginiDNA Found in Foodemmanuel333100% (1)
- Biology Group 2 NotesDocument116 paginiBiology Group 2 NotesAllacheruvu Venkata sai kumarÎncă nu există evaluări
- Bio 101 General BiologyDocument7 paginiBio 101 General BiologyMohammed kashimÎncă nu există evaluări
- Biochemistry Chapter 1 and 2Document120 paginiBiochemistry Chapter 1 and 2Fumofu_2Încă nu există evaluări
- SHS STEM Bio1 Q1 Week 2 Module 3 - Prokaryotic Vs Eukaryotic CellDocument21 paginiSHS STEM Bio1 Q1 Week 2 Module 3 - Prokaryotic Vs Eukaryotic CellEmer Perez100% (7)
- Cells and Cell StructureDocument3 paginiCells and Cell Structureapi-238197463Încă nu există evaluări
- 2ND Quarter Science 7 - Cell Discovery and TheoryDocument3 pagini2ND Quarter Science 7 - Cell Discovery and TheoryLudi Jane TorrefrancaÎncă nu există evaluări
- Krauses Essential HumanDocument315 paginiKrauses Essential HumanEllah MaeÎncă nu există evaluări
- CELLULAR STRUCTURE MICROSCOPYDocument75 paginiCELLULAR STRUCTURE MICROSCOPYfilchibuffÎncă nu există evaluări
- 5090 Scheme of Work (For Examination From 2023)Document80 pagini5090 Scheme of Work (For Examination From 2023)Dipanjana DattaÎncă nu există evaluări
- Motor Proteins Practice Q's AnswersDocument11 paginiMotor Proteins Practice Q's AnswersBlaÎncă nu există evaluări
- Cell Organelles and FunctionsDocument6 paginiCell Organelles and FunctionsMia NavarroÎncă nu există evaluări
- Worksheet No. 1 - Cell OrganelleDocument3 paginiWorksheet No. 1 - Cell OrganelleLaureen BarbsÎncă nu există evaluări
- Diffusion DetailedDocument7 paginiDiffusion Detailedkimberley StuartÎncă nu există evaluări
- Everett Gabel - Activity - Prokaryote and Eukaryote Cells (PG) .PDF - KamiDocument6 paginiEverett Gabel - Activity - Prokaryote and Eukaryote Cells (PG) .PDF - KamiEverett GabelÎncă nu există evaluări
- Modular Worksheet FinalDocument66 paginiModular Worksheet FinalJouichiro Masamune100% (3)
- Organisation of The Organism (Multiple Choice) 1 QPDocument16 paginiOrganisation of The Organism (Multiple Choice) 1 QPNatalie RossetteÎncă nu există evaluări
- Biochemistry ReviewerDocument13 paginiBiochemistry ReviewerGienelle Susana San Juan FerrerÎncă nu există evaluări
- 11 STEM General Biology 1 FIRST QUARTER LMsDocument10 pagini11 STEM General Biology 1 FIRST QUARTER LMsNeil Trezley Sunico Balajadia50% (2)
- M-Caps-01: Botany: NEET - XI StudyingDocument3 paginiM-Caps-01: Botany: NEET - XI StudyingAlokSinghÎncă nu există evaluări
- PHCG 201 CytologyDocument113 paginiPHCG 201 CytologyIbn HaarūnÎncă nu există evaluări
- Prokaryotic and Eukaryotic CellsDocument9 paginiProkaryotic and Eukaryotic Cellsapi-176402481Încă nu există evaluări
- Science 7 w4Document5 paginiScience 7 w4fitz zamoraÎncă nu există evaluări
- Ark Elvin Academy Year 9 Science Study Pack Autumn AssessmentDocument25 paginiArk Elvin Academy Year 9 Science Study Pack Autumn AssessmentLabeenaÎncă nu există evaluări
- Cell Organelles and Their FunctionsDocument3 paginiCell Organelles and Their FunctionsKritzel Mae CastilloÎncă nu există evaluări
- Fucus VesiculosusDocument15 paginiFucus VesiculosusRistaÎncă nu există evaluări
- Biology-Scheme-Of-Work EDEXCELDocument83 paginiBiology-Scheme-Of-Work EDEXCELKim GuermacheÎncă nu există evaluări
- Gsp105 Past Questions&answers-1Document32 paginiGsp105 Past Questions&answers-1JulietÎncă nu există evaluări
- Levy Mwanawasa Medical University Biology AssignmentDocument5 paginiLevy Mwanawasa Medical University Biology AssignmentdianaÎncă nu există evaluări