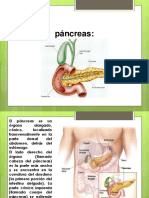
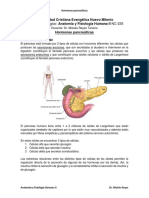
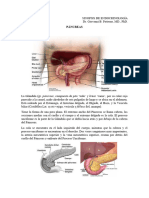
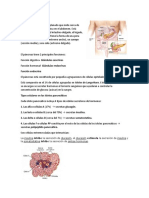
S-ar putea să vă placă și
- Resumen-Fisiología de La Secreción PancreáticaDocument3 paginiResumen-Fisiología de La Secreción PancreáticaJose Alfredo HernandezÎncă nu există evaluări
- Atencion de Enfermería A Pacientes Con Alteraciones EndocrinasDocument30 paginiAtencion de Enfermería A Pacientes Con Alteraciones EndocrinasAgny MoraÎncă nu există evaluări
- MMMMDocument5 paginiMMMMYarib Sarait VillarroelÎncă nu există evaluări
- Sistema Endocrino PancreasDocument7 paginiSistema Endocrino PancreasPatricia DiazÎncă nu există evaluări
- Sistema EndocrinoDocument7 paginiSistema EndocrinoDíana Laura Salinas VillegasÎncă nu există evaluări
- PancreasDocument5 paginiPancreasDayana BerríosÎncă nu există evaluări
- IntroducciónDocument6 paginiIntroducciónYarib Sarait VillarroelÎncă nu există evaluări
- PancreasDocument108 paginiPancreasL- runÎncă nu există evaluări
- El PancreasDocument4 paginiEl PancreasStefani HerreraÎncă nu există evaluări
- PancreasDocument4 paginiPancreasyenniffer yamileth Hodgson LemusÎncă nu există evaluări
- Ana Fisio PancreasDocument9 paginiAna Fisio PancreasAnny EscalonaÎncă nu există evaluări
- Islotes PancreáticosDocument5 paginiIslotes PancreáticosMaría J. Delgado.Încă nu există evaluări
- Presentacion Pancreas, Analisis ClinicosDocument14 paginiPresentacion Pancreas, Analisis ClinicosAndrea De LeonÎncă nu există evaluări
- Fisiologia de Pancreas Endroquino RaipDocument5 paginiFisiologia de Pancreas Endroquino RaipRicardo Iñiguez ParraÎncă nu există evaluări
- PancreasDocument10 paginiPancreasgiuliana fernandez campodonicoÎncă nu există evaluări
- Pancreas NOTAS MEDICASDocument2 paginiPancreas NOTAS MEDICASLuis ValdezÎncă nu există evaluări
- El Sistema PortaDocument6 paginiEl Sistema PortaBeatrizMartinezÎncă nu există evaluări
- DiabetesDocument57 paginiDiabetesSHEYSHEY1993Încă nu există evaluări
- Hormonas y Enzimas PancreticasDocument4 paginiHormonas y Enzimas PancreticasChávez Cruz Sugey YaninÎncă nu există evaluări
- Introducción PancreasDocument7 paginiIntroducción Pancreasdulce miel dulce mielÎncă nu există evaluări
- Páncreas v3Document7 paginiPáncreas v3andrey1232Încă nu există evaluări
- Fisiología Resumen - Regulación Del Sistema DigestivoDocument5 paginiFisiología Resumen - Regulación Del Sistema DigestivoCristhián Ignacio Jerez Fernández100% (1)
- Anatomía Fisiológica Del PancreasDocument4 paginiAnatomía Fisiológica Del PancreasurpiÎncă nu există evaluări
- Secreción PancreáticaDocument3 paginiSecreción PancreáticaSAMIÎncă nu există evaluări
- PancreasDocument203 paginiPancreasMary Salas Maria Salas Ortega100% (3)
- Control de La Secrecion PancreaticaDocument8 paginiControl de La Secrecion PancreaticaSoany M. Alanoca CoaquiraÎncă nu există evaluări
- ENDOCRINOLOGÍA 5ta CLASEDocument17 paginiENDOCRINOLOGÍA 5ta CLASEFlorineÎncă nu există evaluări
- CONCLUSIONESDocument3 paginiCONCLUSIONESRuben LaraÎncă nu există evaluări
- Morfologia Humana - Función de Las Hormonas PancreáticasDocument4 paginiMorfologia Humana - Función de Las Hormonas PancreáticasJunior GuerraÎncă nu există evaluări
- Antiulcerosos y Antidiarreicos CompletDocument44 paginiAntiulcerosos y Antidiarreicos Completflor tellez100% (3)
- Fisiología Del PáncreasDocument3 paginiFisiología Del PáncreasLucia TorresÎncă nu există evaluări
- Glucosa 1Document3 paginiGlucosa 1julieta pavezÎncă nu există evaluări
- BQ Presentación GLUCAGÓN E INSULINADocument39 paginiBQ Presentación GLUCAGÓN E INSULINAmavccÎncă nu există evaluări
- Trabajo IntegradorDocument4 paginiTrabajo IntegradorAbbi NinaÎncă nu există evaluări
- Fisiología DEL PÁNCREASDocument6 paginiFisiología DEL PÁNCREASMarjhorie CaraguayÎncă nu există evaluări
- Cirugía de PáncreasDocument70 paginiCirugía de PáncreasBoby NavarroÎncă nu există evaluări
- Pancreas HormonasDocument25 paginiPancreas Hormonasmcm970380Încă nu există evaluări
- Páncreas Endocrino y Glándula SuprarrenalDocument8 paginiPáncreas Endocrino y Glándula SuprarrenalKadelin Arelis De la RosaÎncă nu există evaluări
- Páncreas EndocrinoDocument3 paginiPáncreas EndocrinoVadi ChÎncă nu există evaluări
- Diabetes Anatomia IntroduccionDocument23 paginiDiabetes Anatomia IntroduccionAlejandro Zamarripa MartínezÎncă nu există evaluări
- PancreasDocument31 paginiPancreasFlor FranchettoÎncă nu există evaluări
- Secrecion Exocrina Del PancreasDocument4 paginiSecrecion Exocrina Del PancreasleilaÎncă nu există evaluări
- Fisiología Del PáncreasDocument6 paginiFisiología Del PáncreasRaúl R. CeledónÎncă nu există evaluări
- PANCREATITISDocument10 paginiPANCREATITISMichelle AvalosÎncă nu există evaluări
- Guía Control Homeostatico GlucosaDocument11 paginiGuía Control Homeostatico GlucosaMaria Ignacia Gaete GodoyÎncă nu există evaluări
- Resumen Modulo2Document11 paginiResumen Modulo2Benjamin Torres GomezÎncă nu există evaluări
- Alteraciones PancreticasDocument8 paginiAlteraciones PancreticasjulissaÎncă nu există evaluări
- Taller Fisiología Páncreas EndocrinoDocument16 paginiTaller Fisiología Páncreas Endocrinopaulina mezaÎncă nu există evaluări
- Células Del PáncreasDocument2 paginiCélulas Del PáncreasLupita NegronÎncă nu există evaluări
- Anatomía Del PáncreasDocument6 paginiAnatomía Del PáncreasD͎i͎e͎g͎o͎ A͎l͎e͎j͎a͎n͎d͎r͎o͎Încă nu există evaluări
- Seminario Tp8 Física BiológicaDocument13 paginiSeminario Tp8 Física BiológicaEstefanía MansillaÎncă nu există evaluări
- VETERINARIA Pancreatitis en Perros y Gatos IDocument10 paginiVETERINARIA Pancreatitis en Perros y Gatos IDa Menra67% (3)
- Fisiologia 100225164223 Phpapp02Document15 paginiFisiologia 100225164223 Phpapp02Laura E. R. ContrerasÎncă nu există evaluări
- Insulina Glucagon y Diabetes Mellitus Capitulo 79 de Fisiologia Humana de Guyton y Hall 13ed 130188 DownloableDocument16 paginiInsulina Glucagon y Diabetes Mellitus Capitulo 79 de Fisiologia Humana de Guyton y Hall 13ed 130188 DownloablePaola Sernaque VilchezÎncă nu există evaluări
- Celula MioDocument10 paginiCelula Mionatalia_cataldoÎncă nu există evaluări
- Anatomia Del Pancreas y FuncionesDocument25 paginiAnatomia Del Pancreas y FuncionesTitulación IFICEDÎncă nu există evaluări
- Ilovepdf MergedDocument78 paginiIlovepdf MergedGabriel EspinoÎncă nu există evaluări
- La Digestión y La CirculaciónDocument4 paginiLa Digestión y La CirculaciónJose Luis Llamoza Romero50% (2)
- La Ciencia Del Metabolismo: Metabolismo sin rodeosDe la EverandLa Ciencia Del Metabolismo: Metabolismo sin rodeosEvaluare: 5 din 5 stele5/5 (1)
- PC1 - PB2 OdontoDocument9 paginiPC1 - PB2 OdontoFlavia RoncalÎncă nu există evaluări
- Cetogeenicants-13-02896 en EsDocument16 paginiCetogeenicants-13-02896 en EsCamilo GarciaÎncă nu există evaluări
- Dieta AnabolicaDocument7 paginiDieta AnabolicaRodrigo Patricio Alegre AlayÎncă nu există evaluări
- 2013 Lopez Morales Edorexia Definicion y VariablesDocument159 pagini2013 Lopez Morales Edorexia Definicion y VariablesPamela BalmoriÎncă nu există evaluări
- DisacáridosDocument11 paginiDisacáridosnhicollÎncă nu există evaluări
- Los Sistemas Energéticos en Las Artes MarcialesDocument3 paginiLos Sistemas Energéticos en Las Artes MarcialesRodrigo ZarateÎncă nu există evaluări
- 1.metabolismo de GlucidosDocument13 pagini1.metabolismo de GlucidosjogugaÎncă nu există evaluări
- CARBOHIDRATOSDocument18 paginiCARBOHIDRATOSMariana ValenciaÎncă nu există evaluări
- Adecuacion FisicaDocument25 paginiAdecuacion FisicaCarlos A. MontañoÎncă nu există evaluări
- Tarea 1 BioquímicaDocument3 paginiTarea 1 BioquímicaAlma Guadalupe Marín MagañaÎncă nu există evaluări
- CARBOHIDRATOSDocument63 paginiCARBOHIDRATOScamila gutierrezÎncă nu există evaluări
- El AMP CíclicoDocument6 paginiEl AMP CíclicoCeci MartinezÎncă nu există evaluări
- Bme U3 Ea VesrDocument6 paginiBme U3 Ea VesrMonse Sanchez100% (1)
- MergedDocument8 paginiMergedCLAUDIA ORTUETAÎncă nu există evaluări
- AMPK y mTORDocument13 paginiAMPK y mTORJorge Alexander Román MoralesÎncă nu există evaluări
- Bioquimica 2 BloqueDocument33 paginiBioquimica 2 BloqueIss DuuarteeÎncă nu există evaluări
- Anaerobiosis MuscularDocument8 paginiAnaerobiosis MuscularMarcelo Anibal Alvarez100% (1)
- Bioquimica Glucogenolisis y GluconeogenesisDocument53 paginiBioquimica Glucogenolisis y GluconeogenesisDaniel JLÎncă nu există evaluări
- Transformación Bioquímica Del Músculo en Carne (Autoguardado)Document22 paginiTransformación Bioquímica Del Músculo en Carne (Autoguardado)Verónica ArévaloÎncă nu există evaluări
- Glicólisis FetalDocument12 paginiGlicólisis FetalPreciado09Încă nu există evaluări
- GlucogenolisisDocument46 paginiGlucogenolisisAnonymous 7y5lhH0100% (1)
- POLISACÁRIDOSDocument55 paginiPOLISACÁRIDOSPiiyyiissÎncă nu există evaluări
- Fisiología Del AyunoDocument7 paginiFisiología Del AyunoGabriela Sofia Cespedes HervasÎncă nu există evaluări
- Metabolismo y NutricionDocument78 paginiMetabolismo y NutricionBrenda MerazÎncă nu există evaluări
- Carbohidratos Hoja de TrabajoDocument5 paginiCarbohidratos Hoja de TrabajoMarbellaÎncă nu există evaluări
- 2023 Sistema EndocrinoDocument64 pagini2023 Sistema EndocrinoDeyerlin MendozaÎncă nu există evaluări
- Diabetes Gestacional (Parte 1)Document13 paginiDiabetes Gestacional (Parte 1)Cristian Velasquez DiazÎncă nu există evaluări
- CarbohidratosDocument86 paginiCarbohidratosJohn Andrés Rodríguez VasquezÎncă nu există evaluări
- Bioquimica II Cfi-235Document56 paginiBioquimica II Cfi-235Lizy MorelÎncă nu există evaluări
- Sesión 3-03 La Circulación y La Excreción en Los Animales 2023Document5 paginiSesión 3-03 La Circulación y La Excreción en Los Animales 2023Fabio CallataÎncă nu există evaluări