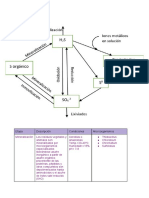
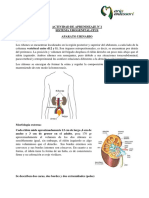
S-ar putea să vă placă și
- Trabajo de Investigación Biofiltros de AlgasDocument7 paginiTrabajo de Investigación Biofiltros de AlgasPauPauSánchezÎncă nu există evaluări
- Práctica #6 FisiologíaDocument14 paginiPráctica #6 FisiologíaPauPauSánchezÎncă nu există evaluări
- Bitacora - Aislamiento - Hidrolisis DNADocument4 paginiBitacora - Aislamiento - Hidrolisis DNAPauPauSánchezÎncă nu există evaluări
- Bitacora Obtención de DNA PlasmídicoDocument3 paginiBitacora Obtención de DNA PlasmídicoPauPauSánchezÎncă nu există evaluări
- 1 4-HidroponiaDocument26 pagini1 4-HidroponiaPauPauSánchezÎncă nu există evaluări
- Manejo de Áreas Bajo La Curva y Tiempos de Retención.Document2 paginiManejo de Áreas Bajo La Curva y Tiempos de Retención.PauPauSánchezÎncă nu există evaluări
- Reporte SEPARACIÓN DE GRUPOS USANDO CROMATOGRAFÍA POR EXCLUSIÓNDocument5 paginiReporte SEPARACIÓN DE GRUPOS USANDO CROMATOGRAFÍA POR EXCLUSIÓNPauPauSánchezÎncă nu există evaluări
- Efecto de La Luz Sobre Las PlantasDocument25 paginiEfecto de La Luz Sobre Las PlantasPauPauSánchezÎncă nu există evaluări
- Biotec-Cuestionario 2Document3 paginiBiotec-Cuestionario 2PauPauSánchezÎncă nu există evaluări
- Exposición SN1 ParcialDocument27 paginiExposición SN1 ParcialPauPauSánchezÎncă nu există evaluări
- Seminario Eco P5.Document34 paginiSeminario Eco P5.PauPauSánchez100% (1)
- TemperaturaDocument22 paginiTemperaturaPauPauSánchezÎncă nu există evaluări
- Protocolo MetodologíaDocument11 paginiProtocolo MetodologíaPauPauSánchezÎncă nu există evaluări
- Sánchez Munguía Ciclos S y NDocument5 paginiSánchez Munguía Ciclos S y NPauPauSánchezÎncă nu există evaluări
- Seminario Eco P5.Document34 paginiSeminario Eco P5.PauPauSánchez100% (1)
- Paulina Sánchez - Ciclos Del Hierro y FósforoDocument4 paginiPaulina Sánchez - Ciclos Del Hierro y FósforoPauPauSánchezÎncă nu există evaluări
- Sánchez Munguía Ciclos S y NDocument5 paginiSánchez Munguía Ciclos S y NPauPauSánchezÎncă nu există evaluări
- Biotec-Cuestionario 1Document3 paginiBiotec-Cuestionario 1PauPauSánchezÎncă nu există evaluări
- Practica 4 y 5 FISIOLOGIADocument26 paginiPractica 4 y 5 FISIOLOGIAPauPauSánchezÎncă nu există evaluări
- Protocolo MetodologíaDocument11 paginiProtocolo MetodologíaPauPauSánchezÎncă nu există evaluări
- PRÁCTICA No 3 PDFDocument5 paginiPRÁCTICA No 3 PDFPauPauSánchezÎncă nu există evaluări
- Practica 4 y 5 FISIOLOGIADocument26 paginiPractica 4 y 5 FISIOLOGIAPauPauSánchezÎncă nu există evaluări
- Evaluacion de Procesos Microbianos en El Ciclo Del CarbonoDocument9 paginiEvaluacion de Procesos Microbianos en El Ciclo Del CarbonoPauPauSánchezÎncă nu există evaluări
- Eco 16 (2) 05Document6 paginiEco 16 (2) 05Tania Xicoténcatl RodríguezÎncă nu există evaluări
- Analisis Especiales Hortalizas PDFDocument38 paginiAnalisis Especiales Hortalizas PDFPauPauSánchezÎncă nu există evaluări
- AMINOACIDOS DIGESTIBLES en PollosDocument19 paginiAMINOACIDOS DIGESTIBLES en PollosManuel Ruiz ZebaduaÎncă nu există evaluări
- Caninos. Retenidos Traccion y Retraccion Mayo 2012Document51 paginiCaninos. Retenidos Traccion y Retraccion Mayo 2012Marisol Ponce De León100% (2)
- INFORME 02 - Genetica VegDocument18 paginiINFORME 02 - Genetica VegRenzo TarrilloÎncă nu există evaluări
- Signos VitalesDocument43 paginiSignos VitalesPierre Alberto Ponte MendozaÎncă nu există evaluări
- Evita Las Noticias - Rolf DobelliDocument9 paginiEvita Las Noticias - Rolf DobellixurxofÎncă nu există evaluări
- Clasificacion de Las EspeciesDocument9 paginiClasificacion de Las Especiesk9mkhczxjxÎncă nu există evaluări
- Evaluación Ciencias Naturales 2 ADocument3 paginiEvaluación Ciencias Naturales 2 AClaudia Godoy EspinozaÎncă nu există evaluări
- Jornalización CienciasDocument3 paginiJornalización CienciasRoxana RivasÎncă nu există evaluări
- PaltasDocument22 paginiPaltasMariana CarrascoÎncă nu există evaluări
- ¿Se Puede Determinar El Punto Isoeléctrico de Caseína y Gelatina Por PrecipitaciónDocument1 pagină¿Se Puede Determinar El Punto Isoeléctrico de Caseína y Gelatina Por PrecipitaciónSheila CruzÎncă nu există evaluări
- Atlas de Anatomia CardiacaDocument176 paginiAtlas de Anatomia CardiacaFJ67% (3)
- PorciculturaDocument64 paginiPorciculturaAdrianSilvaPradaÎncă nu există evaluări
- Actividad de Aprendizaje N°1 Fisiopatologia FarmaciaDocument11 paginiActividad de Aprendizaje N°1 Fisiopatologia FarmaciaRosa Maria Pillco CcahuaÎncă nu există evaluări
- Banco de Preguntas Maniobras PreviasDocument8 paginiBanco de Preguntas Maniobras PreviasMilenka MoreiraÎncă nu există evaluări
- Control de PlagasDocument4 paginiControl de Plagasyanet garciaÎncă nu există evaluări
- Explique Cómo y Cuáles Son Las Diferencias en El Procesamiento de Información en Los Cerebros Masculinos y FemeninosDocument2 paginiExplique Cómo y Cuáles Son Las Diferencias en El Procesamiento de Información en Los Cerebros Masculinos y FemeninosJhon JhoseÎncă nu există evaluări
- CLASE VIRTUAL ESCRITA 13 16 de FEBRERO Evolutivo LemsDocument4 paginiCLASE VIRTUAL ESCRITA 13 16 de FEBRERO Evolutivo Lemsfernando mendozaÎncă nu există evaluări
- Bio - 4° - 2021 (Tema 1) - La Nutrición de Las PlantasDocument4 paginiBio - 4° - 2021 (Tema 1) - La Nutrición de Las PlantasANGELICA SALAZARÎncă nu există evaluări
- (RL) 01-21-Muy Interesante MexicoDocument100 pagini(RL) 01-21-Muy Interesante MexicoMartha Alicia Jaén LaraÎncă nu există evaluări
- Gestion AmbientalDocument32 paginiGestion AmbientalMarielaÎncă nu există evaluări
- Mecanismo de Acción HormonalDocument14 paginiMecanismo de Acción HormonalRaychelk AparicioÎncă nu există evaluări
- Examen 2 ParcialDocument6 paginiExamen 2 ParcialmariajoserojasnietoÎncă nu există evaluări
- Pórfidos de CobreDocument46 paginiPórfidos de CobreJuan Sanchez VelezmoroÎncă nu există evaluări
- Tuneles de CompostajeDocument7 paginiTuneles de Compostajemaura ferrerÎncă nu există evaluări
- MoluscosDocument9 paginiMoluscoslaura natalia castellanos ardilaÎncă nu există evaluări
- Documento Matricula UTNDocument1 paginăDocumento Matricula UTNDayana VasquezÎncă nu există evaluări
- Tema 2 PDFDocument20 paginiTema 2 PDFRocio FreireÎncă nu există evaluări
- Pasos para La Diseccion de Un InsectoDocument1 paginăPasos para La Diseccion de Un Insectoahlcides0% (1)
- DiverticulitisDocument43 paginiDiverticulitisRamónArturoRodríguezÎncă nu există evaluări
- Biologia Celular Veterinaria PDFDocument8 paginiBiologia Celular Veterinaria PDFAli Sobalvarro FischerÎncă nu există evaluări