S-ar putea să vă placă și
- Los Seres Vivos Dependen De La Mecánica CuánticaDe la EverandLos Seres Vivos Dependen De La Mecánica CuánticaÎncă nu există evaluări
- Receptores FarmacológicosDocument13 paginiReceptores FarmacológicosCinthia Diaz100% (1)
- Tarea 3Document4 paginiTarea 3yeseniaÎncă nu există evaluări
- FarmacologiaDocument98 paginiFarmacologiaHECTOR CASTELLANOSÎncă nu există evaluări
- Receptores de FármacosDocument22 paginiReceptores de FármacosRukia SakataÎncă nu există evaluări
- Deber Farmacologia Factores FarmacologicosDocument7 paginiDeber Farmacologia Factores FarmacologicosErika Maribel BadilloÎncă nu există evaluări
- Mecanismos Generales de Acción de Los FármacosDocument6 paginiMecanismos Generales de Acción de Los Fármacosarlekin01Încă nu există evaluări
- Farmaco ParcialDocument17 paginiFarmaco ParcialMilagrosMaidanaÎncă nu există evaluări
- Seminario - Ciclo de Vida de Los Receptores FarmacologicosDocument22 paginiSeminario - Ciclo de Vida de Los Receptores Farmacologicostoch tikÎncă nu există evaluări
- Clasificacion Segun Afinidad Con Receptores de FarmacoterapiaDocument13 paginiClasificacion Segun Afinidad Con Receptores de FarmacoterapiacatsÎncă nu există evaluări
- Criterios de Los Receptores CelularesDocument5 paginiCriterios de Los Receptores CelularesKrolina BeltranÎncă nu există evaluări
- Mecanismos de Accion de Fármacos:: Acción y Efecto FarmacológicosDocument8 paginiMecanismos de Accion de Fármacos:: Acción y Efecto FarmacológicosGustavo RinconÎncă nu există evaluări
- Mecanismo de Acción de Los FármacosDocument7 paginiMecanismo de Acción de Los FármacosMay MozoÎncă nu există evaluări
- Principios de Farmacología MolecularDocument33 paginiPrincipios de Farmacología MolecularLeonelito MedinaÎncă nu există evaluări
- 2) FarmacodinamiaDocument13 pagini2) FarmacodinamiaMilagros SilvaÎncă nu există evaluări
- Nicol Mercedes, Tarea 3Document13 paginiNicol Mercedes, Tarea 3alejandra alcantaraÎncă nu există evaluări
- Resumen Capitulo 4Document5 paginiResumen Capitulo 4Magally Castañeda GómezÎncă nu există evaluări
- Receptores FarmacologíaDocument30 paginiReceptores FarmacologíaBerly Ruiz100% (1)
- Mecanismos de Los FármacosDocument23 paginiMecanismos de Los FármacosPaola Sánchez TapiaÎncă nu există evaluări
- Tipos de Receptores, Pregunta 2 de FarmacodinamiaDocument3 paginiTipos de Receptores, Pregunta 2 de FarmacodinamiaDaniela HernandezÎncă nu există evaluări
- RECEPTORES DE FÁRMACOS - PPSXDocument23 paginiRECEPTORES DE FÁRMACOS - PPSXLuis Barrionuevo QuintanaÎncă nu există evaluări
- Farmacologia MolecularDocument5 paginiFarmacologia MolecularolgakatherineÎncă nu există evaluări
- Monografia Equipo6 FarmacodinamiaDocument2 paginiMonografia Equipo6 FarmacodinamiaTerra NewÎncă nu există evaluări
- Monografia Farmacodinamica - TerminadoDocument14 paginiMonografia Farmacodinamica - TerminadoJimmyÎncă nu există evaluări
- Seminario PPT - Ciclo de Vida de Los ReceptoresDocument13 paginiSeminario PPT - Ciclo de Vida de Los Receptorestoch tikÎncă nu există evaluări
- Mecanismos Moleculares de Acción de Los FármacosDocument8 paginiMecanismos Moleculares de Acción de Los FármacosariarapaÎncă nu există evaluări
- Teoria de ReceptoresDocument2 paginiTeoria de ReceptoresGraciela ReÎncă nu există evaluări
- Guia Desarrollada de Autoaprendizaje 2 Farmacodinamia v2Document15 paginiGuia Desarrollada de Autoaprendizaje 2 Farmacodinamia v2Escobar EvelynÎncă nu există evaluări
- 4 Interacciones Frmaco y ReceptorDocument12 pagini4 Interacciones Frmaco y ReceptorEdwin Valarezo PrecillaÎncă nu există evaluări
- Resumen FarmacodinamiaDocument6 paginiResumen FarmacodinamiaNicole RojasÎncă nu există evaluări
- Mecanismos Biológicos de Interacción de Los Fármacos Con Los SistemasDocument13 paginiMecanismos Biológicos de Interacción de Los Fármacos Con Los SistemasGrettaly GiampaÎncă nu există evaluări
- Clase 2 - Principios Generales de La Acción de Los FármacosDocument59 paginiClase 2 - Principios Generales de La Acción de Los FármacosJose AcostaÎncă nu există evaluări
- Farm A Codina MiaDocument12 paginiFarm A Codina MiaEstefanía OrtizÎncă nu există evaluări
- Cap3 Farmacodinamia Malgor ValseciaDocument7 paginiCap3 Farmacodinamia Malgor ValseciaLuis Diego Caballero EspejoÎncă nu există evaluări
- Receptores y Ligandos FarmacologiaDocument4 paginiReceptores y Ligandos FarmacologiaAndrés Avilés EcheverríaÎncă nu există evaluări
- FARMACOLOGIADocument3 paginiFARMACOLOGIAAndrés Avilés EcheverríaÎncă nu există evaluări
- Blancos FarmacologicosDocument22 paginiBlancos FarmacologicossarahimanzoÎncă nu există evaluări
- BANCO Farma Unidad 1. 2024Document6 paginiBANCO Farma Unidad 1. 2024Nicole Gabriela Rufino IbarraÎncă nu există evaluări
- Farmacología MolecularDocument13 paginiFarmacología Molecularsanx19Încă nu există evaluări
- 3.nomenclatura y Clasificacion de Receptores - InteraccionesDocument4 pagini3.nomenclatura y Clasificacion de Receptores - InteraccionesLion LeoninoÎncă nu există evaluări
- Tema 3 Receptores de FármacosDocument15 paginiTema 3 Receptores de FármacosAlfredo Fernandez GarciaÎncă nu există evaluări
- FarmacodinamiaDocument5 paginiFarmacodinamiaMisael RubioÎncă nu există evaluări
- 02-FARMACODINAMIA ResumenDocument11 pagini02-FARMACODINAMIA ResumenMaxy RosalesÎncă nu există evaluări
- Regulación y DesensibilizaciónDocument12 paginiRegulación y DesensibilizaciónAna Cataño CaicedoÎncă nu există evaluări
- Cuestionario de FarmacoDocument6 paginiCuestionario de FarmacoRolando ValerioÎncă nu există evaluări
- 1.1 RESUMEN-farmacodinamia IDocument19 pagini1.1 RESUMEN-farmacodinamia IAlejandra AlvarezÎncă nu există evaluări
- AgonistasDocument12 paginiAgonistasHabitaciones HospicioÎncă nu există evaluări
- Estela MaryDocument10 paginiEstela MaryOctavio Luis Ramírez DuarteÎncă nu există evaluări
- Receptores Farmacológicos Concepto He HistoriaDocument14 paginiReceptores Farmacológicos Concepto He HistoriaCONCEPCION PEREZ PREZAÎncă nu există evaluări
- DINAMICA 4 (Apuntes)Document2 paginiDINAMICA 4 (Apuntes)Juan FernándezÎncă nu există evaluări
- 4 Diapositiva FarmacodinamiaDocument28 pagini4 Diapositiva FarmacodinamiaYuly Hoyos HidalgoÎncă nu există evaluări
- 6 Metabolismo Celular. Recepción y Traducción de SeñalesDocument10 pagini6 Metabolismo Celular. Recepción y Traducción de Señaleslizbeth TibanlomboÎncă nu există evaluări
- Farmacología USP 6 FinalDocument102 paginiFarmacología USP 6 FinalRay A RmirzÎncă nu există evaluări
- Receptores EndocrinosDocument5 paginiReceptores EndocrinosNayely ErasÎncă nu există evaluări
- Material de Farmacología Segundo ParcialDocument32 paginiMaterial de Farmacología Segundo ParcialXadvier Rivera Jimenez100% (2)
- Receptores FarmacológicosDocument5 paginiReceptores FarmacológicosIvan PonzoÎncă nu există evaluări
- Interacción Fármaco-ReceptorDocument17 paginiInteracción Fármaco-ReceptorJuan Carlos Amaro Bautista100% (1)
- Farmacologia Clase 01Document72 paginiFarmacologia Clase 01Daniel BffaerÎncă nu există evaluări
- Examencarbonilos2019 2 PDFDocument6 paginiExamencarbonilos2019 2 PDFkarenÎncă nu există evaluări
- Artículo 2 ReceptoresDocument12 paginiArtículo 2 ReceptoreskarenÎncă nu există evaluări
- Qué Hace La Química para Obtener Un Hidrocarburo A Partir de OtroDocument3 paginiQué Hace La Química para Obtener Un Hidrocarburo A Partir de OtrokarenÎncă nu există evaluări
- Artículo.1. Dósis Efectiva 50 PDFDocument10 paginiArtículo.1. Dósis Efectiva 50 PDFkarenÎncă nu există evaluări
- Farmacologia de Los Agonistas y Antagonistas de Los Receptores OpiodesDocument32 paginiFarmacologia de Los Agonistas y Antagonistas de Los Receptores OpiodesNaya Nieves RivÎncă nu există evaluări
- Medios de CultivoDocument12 paginiMedios de CultivoLaila Nicol CalderonÎncă nu există evaluări
- Artículo.1. Dósis Efectiva 50 PDFDocument10 paginiArtículo.1. Dósis Efectiva 50 PDFkarenÎncă nu există evaluări
- Poemas Al Estilo ForenseDocument1 paginăPoemas Al Estilo ForensekarenÎncă nu există evaluări
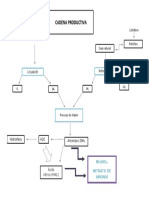
- Cadena Productiva Del Nitrato de AmonioDocument1 paginăCadena Productiva Del Nitrato de AmoniokarenÎncă nu există evaluări
- Cómo Se Separa El PetróleoDocument2 paginiCómo Se Separa El PetróleokarenÎncă nu există evaluări
- Soraya Jiménez (Biografía)Document2 paginiSoraya Jiménez (Biografía)karenÎncă nu există evaluări
- Ser y AcontecerDocument1 paginăSer y AcontecerkarenÎncă nu există evaluări
- Soraya Jiménez (Biografía)Document2 paginiSoraya Jiménez (Biografía)karenÎncă nu există evaluări
- ARTÍC Viruela PDFDocument14 paginiARTÍC Viruela PDFkarenÎncă nu există evaluări
- Articulo EmbarazoDocument23 paginiArticulo EmbarazokarenÎncă nu există evaluări
- Cómo Se Separa El PetróleoDocument2 paginiCómo Se Separa El PetróleokarenÎncă nu există evaluări
- Titulaciones ConductimetricasDocument17 paginiTitulaciones ConductimetricasAlexis Sánchez100% (1)
- 1521 7Document7 pagini1521 7Scott BranchÎncă nu există evaluări
- Espectrofotómetro de Arreglo de DiodosDocument3 paginiEspectrofotómetro de Arreglo de DiodosAnthonio Motha100% (2)
- Manual de Biofarmacia PDFDocument21 paginiManual de Biofarmacia PDFVivi VelascoÎncă nu există evaluări
- 1521 7Document7 pagini1521 7Scott BranchÎncă nu există evaluări
- Esquemas de Destilación A P ReducidaDocument5 paginiEsquemas de Destilación A P ReducidakarenÎncă nu există evaluări
- AcupunturaDocument10 paginiAcupunturakaren0% (1)
- CUADERNILLOInnovacionDocument66 paginiCUADERNILLOInnovacionNestor MotoloÎncă nu există evaluări
- Articulo EmbarazoDocument23 paginiArticulo EmbarazokarenÎncă nu există evaluări
- EspectrofotometriaDocument3 paginiEspectrofotometriasofia9303Încă nu există evaluări
- Destilación A PRDocument62 paginiDestilación A PRkarenÎncă nu există evaluări
- Error FotometricoDocument5 paginiError FotometricokarenÎncă nu există evaluări
- Ley General de Sociedades MercantilesDocument5 paginiLey General de Sociedades MercantileskarenÎncă nu există evaluări
- Fase 4 Transporte de Sólidos y FluidosDocument11 paginiFase 4 Transporte de Sólidos y FluidosJovany rojano100% (1)
- Taller Semana 3 RealizadoDocument3 paginiTaller Semana 3 RealizadoMishi0897100% (3)
- Guia 4 Tipos de Sustancias Grado 9Document4 paginiGuia 4 Tipos de Sustancias Grado 9Benavides Ortiz Luis ErnestoÎncă nu există evaluări
- "Establecimiento de Un Proceso deDocument59 pagini"Establecimiento de Un Proceso deKatherine Rocio Obeid ManjarrezÎncă nu există evaluări
- Curvas de Valoración Acido BaseDocument10 paginiCurvas de Valoración Acido BaseJesus NMÎncă nu există evaluări
- Resumen de enlaces en el ADN: Puentes de Hidrógeno: Unen a las bases nitrogenadas entre sí. A-T Enlace N-β-glucosídico: Unen al Azúcar en C1 con la Enlaces fosfodiesterDocument3 paginiResumen de enlaces en el ADN: Puentes de Hidrógeno: Unen a las bases nitrogenadas entre sí. A-T Enlace N-β-glucosídico: Unen al Azúcar en C1 con la Enlaces fosfodiesterELSA ALEJANDRA CRUZ CARDENASÎncă nu există evaluări
- Balance de Materia NectarDocument3 paginiBalance de Materia NectarRaysha Mileynn IbañezÎncă nu există evaluări
- Unidad 3 - Bioenergética, Enzimas y MetabolismoDocument33 paginiUnidad 3 - Bioenergética, Enzimas y MetabolismoSergio Franco ViverosÎncă nu există evaluări
- Papel Ecológico Elaborado A Partir de Piedra Caliza y PolímeroDocument12 paginiPapel Ecológico Elaborado A Partir de Piedra Caliza y Polímerokevin rodriguezÎncă nu există evaluări
- Capitulo 2 2.0Document38 paginiCapitulo 2 2.0Efraín CórdovaÎncă nu există evaluări
- Huellas ExperimentoDocument18 paginiHuellas ExperimentoLore Wheelock100% (2)
- 2IBPMT222E5Document18 pagini2IBPMT222E5Emanuel MaloÎncă nu există evaluări
- Lluvia ÁcidaDocument5 paginiLluvia ÁcidaMariana VargasÎncă nu există evaluări
- BASEPIROXILINA Duco PDFDocument2 paginiBASEPIROXILINA Duco PDFnsmlmapc100% (1)
- GUIA 6 Problema 10Document2 paginiGUIA 6 Problema 10cruzsilvaÎncă nu există evaluări
- 3 Clase Pavimentos Unet 2017.1Document35 pagini3 Clase Pavimentos Unet 2017.1Anthony VelazcoÎncă nu există evaluări
- Lampara de Calor RadianteDocument2 paginiLampara de Calor RadianteJimena Lopez CardenasÎncă nu există evaluări
- PROTOCOLO DE APLICACIÓN PARA MEDIR PH EN AGUADocument2 paginiPROTOCOLO DE APLICACIÓN PARA MEDIR PH EN AGUAAlanÎncă nu există evaluări
- MiniproyectoDocument41 paginiMiniproyectoNelson Aron Llacsahuanga HerreraÎncă nu există evaluări
- Trastornos HidroelectrolíticosDocument65 paginiTrastornos HidroelectrolíticosSofi PetroÎncă nu există evaluări
- Instrumental de Laboratorio Bioquimica Sebastian RodriguezDocument4 paginiInstrumental de Laboratorio Bioquimica Sebastian RodriguezSebastian RodriguezÎncă nu există evaluări
- Materias PrimasDocument8 paginiMaterias Primasmorris mendez89% (19)
- T 6. La Célula y Las Envolturas CelularesDocument11 paginiT 6. La Célula y Las Envolturas CelularesIrene García MarcosÎncă nu există evaluări
- Tarea 1 Diseño y Simulacion de Procesos y Productos 06092021Document11 paginiTarea 1 Diseño y Simulacion de Procesos y Productos 06092021Jose OctavioÎncă nu există evaluări
- Reacciones RedoxDocument4 paginiReacciones Redoxyojan fuentesÎncă nu există evaluări
- Fotosíntesis 1Document3 paginiFotosíntesis 1Adelyna LimaÎncă nu există evaluări
- RRamos Nomenclatura AlcoholesDocument10 paginiRRamos Nomenclatura AlcoholesrobertoarnulforamosÎncă nu există evaluări
- Resumen ConstruccionesDocument31 paginiResumen ConstruccionesDavid LugoÎncă nu există evaluări
- Produtos Químicos Orgânicos - Carballido Tomo I Volume IIDocument948 paginiProdutos Químicos Orgânicos - Carballido Tomo I Volume IISonia Regina dos SantosÎncă nu există evaluări
- Lab Cultivo de TejidosDocument32 paginiLab Cultivo de Tejidosa1165990Încă nu există evaluări