S-ar putea să vă placă și
- Nepeta Paper 2021Document10 paginiNepeta Paper 2021Dr. Ali NoorÎncă nu există evaluări
- Spectrophotometric Determination of Adrenaline in Pharmaceutical PreparationsDocument5 paginiSpectrophotometric Determination of Adrenaline in Pharmaceutical PreparationsmayaÎncă nu există evaluări
- IJPPR, Vol 9, Issue 6, Article 3Document5 paginiIJPPR, Vol 9, Issue 6, Article 3yahyaÎncă nu există evaluări
- Identification of Phenolic Acid From Ethanol ExtraDocument10 paginiIdentification of Phenolic Acid From Ethanol ExtraVennyÎncă nu există evaluări
- Rapid Screening and Identification of Antioxidants in Aqueous Ex - 2009 - Food CDocument7 paginiRapid Screening and Identification of Antioxidants in Aqueous Ex - 2009 - Food CTheerapat Chalerm-MuangÎncă nu există evaluări
- UntitledDocument19 paginiUntitledSarbjeet SinghÎncă nu există evaluări
- Tingkatan AntioksidanDocument5 paginiTingkatan AntioksidanMygameÎncă nu există evaluări
- Molecular Evaluation of Herbal Compounds As Potent Inhibitors of Acetylcholinesterase For The Treatment of Alzheimer's DiseaseDocument7 paginiMolecular Evaluation of Herbal Compounds As Potent Inhibitors of Acetylcholinesterase For The Treatment of Alzheimer's Diseaselehongduong1997Încă nu există evaluări
- Makino 2001Document4 paginiMakino 2001aji gumelarÎncă nu există evaluări
- Kandungan Kimia Dan Uji Antiinflamasi Ekstrak Etanol Lantana Camara L Pada Tikus Putih Rattus Norvegicus L Jantan CompressDocument8 paginiKandungan Kimia Dan Uji Antiinflamasi Ekstrak Etanol Lantana Camara L Pada Tikus Putih Rattus Norvegicus L Jantan Compressdian tiniÎncă nu există evaluări
- Antioxidant Activities of Leaf Extract of Salvia Miltiorrhiza BungeDocument7 paginiAntioxidant Activities of Leaf Extract of Salvia Miltiorrhiza BungeAsma SedÎncă nu există evaluări
- Sedative and Cardiovascular Effects of Aloysia Citriodora Palau, On Mice and RatsDocument8 paginiSedative and Cardiovascular Effects of Aloysia Citriodora Palau, On Mice and RatsFabio PicassoÎncă nu există evaluări
- 712-Article Text-2063-1-10-20131115Document2 pagini712-Article Text-2063-1-10-20131115Gek NikenÎncă nu există evaluări
- Antioxidant Activity of Alkaloid Compounds From Litsea Cubeba LourDocument4 paginiAntioxidant Activity of Alkaloid Compounds From Litsea Cubeba LourSony Eka NugrahaÎncă nu există evaluări
- Advpub Ess14061Document10 paginiAdvpub Ess14061Dewi nur thohidahÎncă nu există evaluări
- 484-Article Text-860-1-10-20181031Document9 pagini484-Article Text-860-1-10-20181031Recky PatalaÎncă nu există evaluări
- Plantas Que Ajudam A Controlar o HivDocument5 paginiPlantas Que Ajudam A Controlar o HivIgor FernandesÎncă nu există evaluări
- Carbonara 2012Document8 paginiCarbonara 2012Manuel Rios GonzalesÎncă nu există evaluări
- Ferlemi2015Document11 paginiFerlemi2015josethrasecÎncă nu există evaluări
- Oil of Lavandula Angustifolia On Amyloid Beta PolymerizationDocument7 paginiOil of Lavandula Angustifolia On Amyloid Beta PolymerizationarcherselevatorsÎncă nu există evaluări
- A Comparision of The Effects of Aloe Barbadensis and Allium Albino RatsDocument4 paginiA Comparision of The Effects of Aloe Barbadensis and Allium Albino RatsEric GibsonÎncă nu există evaluări
- NPC Natural Product CommunicationsDocument6 paginiNPC Natural Product CommunicationsAyu DemiÎncă nu există evaluări
- Phytochemical and Pharmacological Investigations of Eclipta Alba (Linn.)Document5 paginiPhytochemical and Pharmacological Investigations of Eclipta Alba (Linn.)Thu TongÎncă nu există evaluări
- Wang (2012)Document3 paginiWang (2012)Ally BÎncă nu există evaluări
- Paper 1Document6 paginiPaper 1Gopika SureshÎncă nu există evaluări
- Inflammation 1Document7 paginiInflammation 1BRAYAN JOSE HERRERA HERNANDEZ ESTUDIANTE ACTIVOÎncă nu există evaluări
- Formulation, Evaluation and Optimization of Floating Matrix Tablets of CarvedilolDocument15 paginiFormulation, Evaluation and Optimization of Floating Matrix Tablets of CarvedilolBaru Chandrasekhar RaoÎncă nu există evaluări
- Qualitative Estimation of The Presence of Bioactive Compound in Centella Asiatica An Important Medicinal PlantDocument3 paginiQualitative Estimation of The Presence of Bioactive Compound in Centella Asiatica An Important Medicinal Plantscientist786Încă nu există evaluări
- 2716 PDFDocument4 pagini2716 PDFSeptya widhiafniÎncă nu există evaluări
- Amornrat san,+Journal+manager,+SMJ+v3+240-245+RattanaDocument6 paginiAmornrat san,+Journal+manager,+SMJ+v3+240-245+Rattananoviantyramadhani12Încă nu există evaluări
- ACE InhibitorDocument6 paginiACE InhibitorLe XoanÎncă nu există evaluări
- QsarDocument13 paginiQsarLeTienDungÎncă nu există evaluări
- In-Vitro Screening For Acetylcholinesterase Enzyme Inhibition PotentialDocument5 paginiIn-Vitro Screening For Acetylcholinesterase Enzyme Inhibition PotentialRony MandalÎncă nu există evaluări
- Antioxidative and in Vitro Antiproliferative Activity of Arctium Lappa Root ExtractsDocument5 paginiAntioxidative and in Vitro Antiproliferative Activity of Arctium Lappa Root ExtractsMargatzyÎncă nu există evaluări
- The Scientific Basis For The Reputed Activity of ValerianDocument8 paginiThe Scientific Basis For The Reputed Activity of ValerianDiego Bedón AscurraÎncă nu există evaluări
- Hohmann 2011Document6 paginiHohmann 2011aji gumelarÎncă nu există evaluări
- Immunomodulatory Activity of Chlorophytum Borivilianum Sant. FDocument6 paginiImmunomodulatory Activity of Chlorophytum Borivilianum Sant. FNober Sandy LayukÎncă nu există evaluări
- AVOvasorelaxantDocument7 paginiAVOvasorelaxantKate RosalesÎncă nu există evaluări
- Anti-Inflammatory and Analgesic Activities of Ethyl Acetate and Petroleum Ether Fractions of Cassia Auriculata Linn. LeavesDocument7 paginiAnti-Inflammatory and Analgesic Activities of Ethyl Acetate and Petroleum Ether Fractions of Cassia Auriculata Linn. LeavesAsmita MaliÎncă nu există evaluări
- 036 PandeDocument6 pagini036 PandeabdurrahmanassudaisÎncă nu există evaluări
- Role of Hydrogen Bonding in General Anesthesia - XPHDocument6 paginiRole of Hydrogen Bonding in General Anesthesia - XPHdeaaprÎncă nu există evaluări
- RSCB - Binding Affinity - Paper Number 1Document6 paginiRSCB - Binding Affinity - Paper Number 1Ruswanto RuswantoÎncă nu există evaluări
- Total Phenolic and Flavonoid Contents and Antioxidant Activities of Two Raphanus Sativus L. Cultivars (Cherry Belle and Valentine)Document6 paginiTotal Phenolic and Flavonoid Contents and Antioxidant Activities of Two Raphanus Sativus L. Cultivars (Cherry Belle and Valentine)Almas TÎncă nu există evaluări
- 10 1002@pca 1119Document9 pagini10 1002@pca 1119venilia makatambaÎncă nu există evaluări
- Alkal Z Chinskich Ziol '11Document5 paginiAlkal Z Chinskich Ziol '11Paulina CiurykÎncă nu există evaluări
- AntibacterialDocument50 paginiAntibacterial17O1O85 Mutiara Nurul Aulia FirdausÎncă nu există evaluări
- Aronia Acta Med.Document8 paginiAronia Acta Med.Jelena MrmošaninÎncă nu există evaluări
- Lanvers Et Al 2002Document10 paginiLanvers Et Al 2002gemaÎncă nu există evaluări
- Immunomodulator Activity PDFDocument3 paginiImmunomodulator Activity PDFGerald ParkerÎncă nu există evaluări
- IMRAD ParagisDocument11 paginiIMRAD ParagisGaebrelle BoiserÎncă nu există evaluări
- DPPH Free Radical Scavenging Activity of Two Extracts From Agelanthus Dodoneifolius (Loranthaceae) LeavesDocument4 paginiDPPH Free Radical Scavenging Activity of Two Extracts From Agelanthus Dodoneifolius (Loranthaceae) LeavesbryanÎncă nu există evaluări
- JurnalDocument5 paginiJurnalfitrhaÎncă nu există evaluări
- Phillipson, Anderson - 1989 - Ethnopharmacology and Western MedicineDocument12 paginiPhillipson, Anderson - 1989 - Ethnopharmacology and Western MedicineIntan Nabila Sufi ZikrinaÎncă nu există evaluări
- MainDocument7 paginiMainLeena John MathaiÎncă nu există evaluări
- Chemometric Analysis of Selected Medicinal Plants From SerbiaDocument11 paginiChemometric Analysis of Selected Medicinal Plants From SerbiagotcanÎncă nu există evaluări
- Natural ProductsDocument6 paginiNatural ProductsRohan PalÎncă nu există evaluări
- Aegle FruitDocument6 paginiAegle FruitAmmar MasroorÎncă nu există evaluări
- ALZHEIMERDocument10 paginiALZHEIMERalbertompgÎncă nu există evaluări
- 1 ExtractionDocument5 pagini1 ExtractionJoshua FerrerÎncă nu există evaluări
- LC Determination in Sac Intestine Model PDFDocument12 paginiLC Determination in Sac Intestine Model PDFJose PerezÎncă nu există evaluări
- Navarrete Et Al J AOAC International 89 (1) 1-7 (2006) PDFDocument7 paginiNavarrete Et Al J AOAC International 89 (1) 1-7 (2006) PDFJose PerezÎncă nu există evaluări
- P Rotection Against UVB Irradiation by Natural Filters Extracted From Lichens PDFDocument7 paginiP Rotection Against UVB Irradiation by Natural Filters Extracted From Lichens PDFJose PerezÎncă nu există evaluări
- Effect of SKF-525A On Liver Metabolism and Hepatotoxicity of Tri-And Dibutyltin Compounds in MiceDocument6 paginiEffect of SKF-525A On Liver Metabolism and Hepatotoxicity of Tri-And Dibutyltin Compounds in MiceJose PerezÎncă nu există evaluări
- Castillo Juarez Helycobacter PyloriDocument4 paginiCastillo Juarez Helycobacter PyloriJose PerezÎncă nu există evaluări
- Ampphitergium Adstringens PDFDocument10 paginiAmpphitergium Adstringens PDFJose PerezÎncă nu există evaluări
- Drug Absorption Gastrointestinal TractDocument23 paginiDrug Absorption Gastrointestinal TractJose PerezÎncă nu există evaluări
- Nihms 123081Document15 paginiNihms 123081Jose PerezÎncă nu există evaluări
- AnticancerMexicanplants PDFDocument28 paginiAnticancerMexicanplants PDFJose PerezÎncă nu există evaluări
- AfricanPharmacopoeia PDFDocument533 paginiAfricanPharmacopoeia PDFJose Perez50% (2)
- MDMB 4en Pinaca Review 2020Document16 paginiMDMB 4en Pinaca Review 2020AlfonsoÎncă nu există evaluări
- Concepts in Clinical Pharmacokinetics 7th Edition PDFDocument295 paginiConcepts in Clinical Pharmacokinetics 7th Edition PDFsaravanan100% (4)
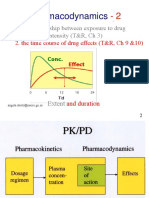
- Introduction and Pharmacodynamics and Drug and Development RegulationDocument91 paginiIntroduction and Pharmacodynamics and Drug and Development RegulationKenneth NuñezÎncă nu există evaluări
- The MOR (Mu Opioid Receptor) and FLNA (Filamin A)Document28 paginiThe MOR (Mu Opioid Receptor) and FLNA (Filamin A)lara gay villaruzÎncă nu există evaluări
- 2021 - BIOM2402 Organ Bath Lab NotesDocument6 pagini2021 - BIOM2402 Organ Bath Lab NotesBlake FangirlÎncă nu există evaluări
- First Sem Pharma NotesDocument75 paginiFirst Sem Pharma NotesLindsay LunodÎncă nu există evaluări
- bmsc13010 Exam t3 2017Document9 paginibmsc13010 Exam t3 2017sandeep kesarÎncă nu există evaluări
- Antioxidant Properties of Cocoa Pods and Shells PDFDocument8 paginiAntioxidant Properties of Cocoa Pods and Shells PDFTài ColeÎncă nu există evaluări
- Nanoclick: A High Throughput, Target-Agnostic Peptide Cell Permeability AssayDocument17 paginiNanoclick: A High Throughput, Target-Agnostic Peptide Cell Permeability AssayMoritz ListÎncă nu există evaluări
- PHARMA 02. PharmacodynamicsDocument11 paginiPHARMA 02. PharmacodynamicsCindy Mae MacamayÎncă nu există evaluări
- Pharmacokinetics Vs PharmacodynamicsDocument41 paginiPharmacokinetics Vs Pharmacodynamicsfarjanaislam mituÎncă nu există evaluări
- Dose Response CurveDocument29 paginiDose Response CurveMaria khurshidÎncă nu există evaluări
- Pharmacology Pharmacodynamics: Dr. Syed Muneeb Anjum (PH.D.) Ips, UvasDocument10 paginiPharmacology Pharmacodynamics: Dr. Syed Muneeb Anjum (PH.D.) Ips, UvasSultan Abdul Hamid IIÎncă nu există evaluări
- Drug Discovery and Development Lecture NotesDocument75 paginiDrug Discovery and Development Lecture NotesJameel BakhshÎncă nu există evaluări
- Pharma S01 SBR03 Le01Document12 paginiPharma S01 SBR03 Le01sky vallartaÎncă nu există evaluări
- KFK200 PD-2 HT22Document40 paginiKFK200 PD-2 HT22Lihle MÎncă nu există evaluări
- USMLE Road Map Pharmacology PDFDocument497 paginiUSMLE Road Map Pharmacology PDFYeshaa Mirani100% (1)
- A Pharmacology Primer 5th EditionDocument7 paginiA Pharmacology Primer 5th Editionarief satyaÎncă nu există evaluări
- IVMS Pharmacology Flash FactsDocument8.250 paginiIVMS Pharmacology Flash FactsMarc Imhotep Cray, M.D.Încă nu există evaluări
- Advances in Therapeutic Peptides Targeting G Protein-Coupled ReceptorsDocument25 paginiAdvances in Therapeutic Peptides Targeting G Protein-Coupled ReceptorsAyberk BinbayÎncă nu există evaluări
- Pharmacology Block 1.3 - PharmacodynamicsDocument12 paginiPharmacology Block 1.3 - Pharmacodynamicsdaleng subÎncă nu există evaluări
- 3,4 Methylenedioxypyrovalerone (MDPV)Document37 pagini3,4 Methylenedioxypyrovalerone (MDPV)Mike RohrichÎncă nu există evaluări
- EC50 ValueDocument8 paginiEC50 ValuePiyush BhallaÎncă nu există evaluări
- 〈121〉 Insulin AssaysDocument6 pagini〈121〉 Insulin AssaysAvijit HazraÎncă nu există evaluări
- Early Drug Discovery ProcessDocument58 paginiEarly Drug Discovery ProcessFrietzyl Mae Generalao100% (1)
- ARTÍCULO LABORATORIO-Quantifying Drug Combination Synergy Along Potency and Efficacy AxesDocument29 paginiARTÍCULO LABORATORIO-Quantifying Drug Combination Synergy Along Potency and Efficacy AxesPame ConzaÎncă nu există evaluări
- Margo - 3 NewDocument71 paginiMargo - 3 NewKhushman KaurÎncă nu există evaluări
- GraphPad Dose Response Ebook PDFDocument12 paginiGraphPad Dose Response Ebook PDFAngeronaÎncă nu există evaluări
- B Udapest University of Technology and Economics Environmental ToxicologyDocument6 paginiB Udapest University of Technology and Economics Environmental ToxicologyArtur Elias FernandesÎncă nu există evaluări
- Analysis of Dose Response Relationship: by Lee Eun JinDocument63 paginiAnalysis of Dose Response Relationship: by Lee Eun JinSunil100% (1)