S-ar putea să vă placă și
- Renal Reflections: Exploring the Marvels of Nephrology: The Intricate Ballet of Kidney Functions UnveiledDe la EverandRenal Reflections: Exploring the Marvels of Nephrology: The Intricate Ballet of Kidney Functions UnveiledÎncă nu există evaluări
- Renal Blood Flow and OxygenationDocument12 paginiRenal Blood Flow and OxygenationnanreÎncă nu există evaluări
- International Review of Experimental Pathology: Kidney DiseaseDe la EverandInternational Review of Experimental Pathology: Kidney DiseaseG. W. RichterEvaluare: 5 din 5 stele5/5 (1)
- Darrow1950 PDFDocument9 paginiDarrow1950 PDFLuís Eduardo CruvinelÎncă nu există evaluări
- Sepsis-Flujo Sanguineo Mesenterico y Shock-2021Document9 paginiSepsis-Flujo Sanguineo Mesenterico y Shock-2021Elsa Pimentel ArceÎncă nu există evaluări
- Hiponatremia PDFDocument11 paginiHiponatremia PDFNessa RojasÎncă nu există evaluări
- BCP 14752Document13 paginiBCP 14752Story WA DokterÎncă nu există evaluări
- Electrolytes and Acid Base Common Fluid and Enlectrolyte DisordersDocument9 paginiElectrolytes and Acid Base Common Fluid and Enlectrolyte Disordersninta karinaÎncă nu există evaluări
- 101 OHara PDFDocument11 pagini101 OHara PDFAntonio Lopez HernandezÎncă nu există evaluări
- Renal Autoregulation and Blood Pressure Management in Circulatory ShockDocument9 paginiRenal Autoregulation and Blood Pressure Management in Circulatory ShockEdson MarquesÎncă nu există evaluări
- Fluid Management: Craig D. Mcclain and Michael L. McmanusDocument25 paginiFluid Management: Craig D. Mcclain and Michael L. McmanusAzzam SaqrÎncă nu există evaluări
- Renal Metabolism and Hypertension Review ArticleDocument12 paginiRenal Metabolism and Hypertension Review ArticlePATRICIA JACQUELINE SARMIENTO GOMEZÎncă nu există evaluări
- Disorders of Water and Sodium BalanceDocument21 paginiDisorders of Water and Sodium BalanceStanÎncă nu există evaluări
- Review Article: Regulation of Cerebral Blood FlowDocument8 paginiReview Article: Regulation of Cerebral Blood FlowAlmas TÎncă nu există evaluări
- Diuretics: Dr. Ashley M. StokesDocument26 paginiDiuretics: Dr. Ashley M. StokesDhaif dhaifÎncă nu există evaluări
- The Pathophysiology of Acute Renal Failure: D P P P, G P. K K K, A G. B B B, S V. SDocument2 paginiThe Pathophysiology of Acute Renal Failure: D P P P, G P. K K K, A G. B B B, S V. SLOLOÎncă nu există evaluări
- Demystifying HyponatremiaDocument10 paginiDemystifying HyponatremiaJavier Vivanco100% (1)
- Fluid Retention in Cirrhosis: Pathophysiology and ManagementDocument16 paginiFluid Retention in Cirrhosis: Pathophysiology and ManagementÃßD ÔûñîÎncă nu există evaluări
- Circadian Rhythms and The KidneyDocument10 paginiCircadian Rhythms and The KidneyneurojuancarfabÎncă nu există evaluări
- Dehert 2012Document11 paginiDehert 2012putriÎncă nu există evaluări
- Cho 2002Document7 paginiCho 2002DanteÎncă nu există evaluări
- Rose 1Document7 paginiRose 1isa mahendraÎncă nu există evaluări
- Jurnal CardioDocument11 paginiJurnal Cardiomohamad safiiÎncă nu există evaluări
- Rheology For CliniciansDocument8 paginiRheology For CliniciansRaluca LÎncă nu există evaluări
- Liquidos y Electrolitos 2Document4 paginiLiquidos y Electrolitos 2Jorge MBÎncă nu există evaluări
- Ehp Si Hiponatremia 1Document8 paginiEhp Si Hiponatremia 1Oana Cristina PetreaÎncă nu există evaluări
- Nutrients: Hydration Status and Cardiovascular FunctionDocument21 paginiNutrients: Hydration Status and Cardiovascular FunctionS Dian RÎncă nu există evaluări
- ANT 2 S20.fullDocument13 paginiANT 2 S20.fulllguerreroÎncă nu există evaluări
- Articolo 21Document32 paginiArticolo 21Francesca BertaccaÎncă nu există evaluări
- Sodium, Water and Potassium: Michael D. PenneyDocument39 paginiSodium, Water and Potassium: Michael D. PenneyChristianus LeonardÎncă nu există evaluări
- Etiology and Management of EdemaDocument14 paginiEtiology and Management of EdemaFavio Alvarez De BejarÎncă nu există evaluări
- Biomarkers of Volume Overload and Edema in HFDocument14 paginiBiomarkers of Volume Overload and Edema in HFalcasan63Încă nu există evaluări
- Piis0007091217540933 PDFDocument6 paginiPiis0007091217540933 PDFAde RohmanaÎncă nu există evaluări
- Congestive Renal FailureDocument9 paginiCongestive Renal FailureSayf QisthiÎncă nu există evaluări
- Manejo en PediatriaDocument15 paginiManejo en PediatriaAlex CalderónÎncă nu există evaluări
- Yaffe Chapter Renal MaturationDocument17 paginiYaffe Chapter Renal Maturationwalaa alsharanyÎncă nu există evaluări
- Physiology AssignmentDocument7 paginiPhysiology AssignmentIdenyi Daniel Ewa EdeÎncă nu există evaluări
- Evidence-Based Hyponatremia Management in Liver DiseaseDocument21 paginiEvidence-Based Hyponatremia Management in Liver DiseaseDea Fiesta JatikusumaÎncă nu există evaluări
- VET343 Urinary Lecture Notes 2014Document35 paginiVET343 Urinary Lecture Notes 2014walczakcÎncă nu există evaluări
- Fluido y ElectrolitosDocument9 paginiFluido y ElectrolitosHen RyÎncă nu există evaluări
- Anuria Oliguria: A Review RatesDocument7 paginiAnuria Oliguria: A Review Ratessurya rajÎncă nu există evaluări
- Fluid Electrolytes and Nutrition Physiological and Clinical AspectsDocument14 paginiFluid Electrolytes and Nutrition Physiological and Clinical AspectsPaulynhiita FashyOnÎncă nu există evaluări
- Renal Lecture 1 PDFDocument29 paginiRenal Lecture 1 PDFSirajum MuneeraÎncă nu există evaluări
- Venous Return in Critical IllnessDocument15 paginiVenous Return in Critical IllnessInês MendonçaÎncă nu există evaluări
- 12 SeminarioDocument6 pagini12 SeminarioJonathanÎncă nu există evaluări
- Fluid Therapy and The Microcirculation in Health and Critical IllnessDocument11 paginiFluid Therapy and The Microcirculation in Health and Critical IllnessAdote DrmÎncă nu există evaluări
- A New Physiologic Based Integrated Algorithm in The Management of Neonatal Hemodynamic InstabilityDocument15 paginiA New Physiologic Based Integrated Algorithm in The Management of Neonatal Hemodynamic InstabilityzeltzinxiomaraÎncă nu există evaluări
- 1994 UrsinoDocument16 pagini1994 UrsinosebasgoryÎncă nu există evaluări
- Physiology of Perfusion of The Secondary Circulatory System in FishDocument1 paginăPhysiology of Perfusion of The Secondary Circulatory System in FishAlejandraÎncă nu există evaluări
- The Kidney - An Organ of Critical Importance in Physiology: Louise RobsonDocument2 paginiThe Kidney - An Organ of Critical Importance in Physiology: Louise Robsoncahyadi adityaÎncă nu există evaluări
- Renal Autoregulation: New Perspectives Regarding The Protective and Regulatory Roles of The Underlying MechanismsDocument27 paginiRenal Autoregulation: New Perspectives Regarding The Protective and Regulatory Roles of The Underlying MechanismsYoAmoNYCÎncă nu există evaluări
- Robert W. Schrier - Atlas of Diseases of The Kidney Volume 03 PDFDocument175 paginiRobert W. Schrier - Atlas of Diseases of The Kidney Volume 03 PDFRobertoBacaÎncă nu există evaluări
- Anesthetic Concerns in Renal FailureDocument16 paginiAnesthetic Concerns in Renal FailureEdwin Gustavo Sandoval TorresÎncă nu există evaluări
- Dialysis Disequilibrium SyndromeDocument7 paginiDialysis Disequilibrium Syndromesatyagraha84Încă nu există evaluări
- Recent Advances in Blood Rheology A ReviewDocument22 paginiRecent Advances in Blood Rheology A ReviewDarius PavriÎncă nu există evaluări
- Ebp 8 72 PDFDocument10 paginiEbp 8 72 PDFMelati HasnailÎncă nu există evaluări
- General Principles of Disorders of Water Balance (Hyponatremia and Hypernatremia) and Sodium Balance (Hypovolemia and Edema) - UpToDateDocument24 paginiGeneral Principles of Disorders of Water Balance (Hyponatremia and Hypernatremia) and Sodium Balance (Hypovolemia and Edema) - UpToDateNedelcu MirunaÎncă nu există evaluări
- Sir David Cuthbertson Medal Lecture - Fluid, Electrolytes and Nutrition: Physiological and Clinical AspectsDocument15 paginiSir David Cuthbertson Medal Lecture - Fluid, Electrolytes and Nutrition: Physiological and Clinical AspectsThinh VinhÎncă nu există evaluări
- 1 s2.0 S0085253815500719 MainDocument4 pagini1 s2.0 S0085253815500719 Mainvalentina berrocal calderonÎncă nu există evaluări
- Metabolic Markers in Sports MedicineDocument55 paginiMetabolic Markers in Sports MedicineallerasicÎncă nu există evaluări
- Symposium On "100 Years of Vitamins" Past, Present, Future: Micronutrients, Macro ImpactDocument3 paginiSymposium On "100 Years of Vitamins" Past, Present, Future: Micronutrients, Macro ImpactallerasicÎncă nu există evaluări
- Spinal C SomatosensoryDocument38 paginiSpinal C SomatosensoryallerasicÎncă nu există evaluări
- FoodDocument8 paginiFoodallerasicÎncă nu există evaluări
- Topic:Control of Blood Pressure: MD - Jabiur Rahaman Daffodil International University Dept. of PharmacyDocument14 paginiTopic:Control of Blood Pressure: MD - Jabiur Rahaman Daffodil International University Dept. of PharmacyallerasicÎncă nu există evaluări
- Motor Functions of The Spina L Cord The Cord Reflexes: Done by Dr. Nour KasbariDocument39 paginiMotor Functions of The Spina L Cord The Cord Reflexes: Done by Dr. Nour KasbariallerasicÎncă nu există evaluări
- Basal ggl1Document24 paginiBasal ggl1allerasicÎncă nu există evaluări
- 2011 Motor Lecture 5 CortexDocument51 pagini2011 Motor Lecture 5 CortexallerasicÎncă nu există evaluări
- Nagasawa Oxitocin Mother InfantDocument10 paginiNagasawa Oxitocin Mother InfantallerasicÎncă nu există evaluări
- Ormerod-Radial Maze ProtocolDocument14 paginiOrmerod-Radial Maze ProtocolallerasicÎncă nu există evaluări
- Basal Ggl1Document24 paginiBasal Ggl1allerasicÎncă nu există evaluări
- Non Nuclear Actions of EstrogenDocument10 paginiNon Nuclear Actions of EstrogenallerasicÎncă nu există evaluări
- Osmotic PumpDocument8 paginiOsmotic PumpallerasicÎncă nu există evaluări
- Hippocamp MemoryDocument3 paginiHippocamp MemoryallerasicÎncă nu există evaluări
- Potassium BalanceDocument6 paginiPotassium BalanceallerasicÎncă nu există evaluări
- Autonomic Function PrognosisDocument5 paginiAutonomic Function PrognosisallerasicÎncă nu există evaluări
- Conjugated BilirubinDocument3 paginiConjugated BilirubinallerasicÎncă nu există evaluări
- Edexecl IGCSE Biology (9 1) Getting Started GuideDocument23 paginiEdexecl IGCSE Biology (9 1) Getting Started GuideKc Kan0% (1)
- General Biology Q4 M3Document16 paginiGeneral Biology Q4 M3Delfin LeeÎncă nu există evaluări
- Nephrotic Syndrome - Armando HasudunganDocument18 paginiNephrotic Syndrome - Armando HasudunganzahraaÎncă nu există evaluări
- Detailed Test Planner: NEET-XII (Passed)Document37 paginiDetailed Test Planner: NEET-XII (Passed)aryanÎncă nu există evaluări
- SCIT 1408 Applied Human Anatomy and Physiology II - Urinary System Chapter 25 BDocument50 paginiSCIT 1408 Applied Human Anatomy and Physiology II - Urinary System Chapter 25 BChuongÎncă nu există evaluări
- BBM - 978 3 319 23458 8/1 PDFDocument22 paginiBBM - 978 3 319 23458 8/1 PDFmedicalÎncă nu există evaluări
- Pharmacology-I Practical ManualDocument24 paginiPharmacology-I Practical ManualJunaed Ahmed50% (2)
- Kidney Lab KeyDocument7 paginiKidney Lab Keyapi-448873912Încă nu există evaluări
- Renal PhysioDocument140 paginiRenal Physioarturo quevedoÎncă nu există evaluări
- Urinary SystemDocument42 paginiUrinary Systemlil'sookie100% (2)
- Primer: Acute Kidney InjuryDocument17 paginiPrimer: Acute Kidney InjuryDiana AngelesÎncă nu există evaluări
- SJFJFDocument9 paginiSJFJFcharizze alpechiÎncă nu există evaluări
- Anatomy and Physiology - Kidney - ForDocument3 paginiAnatomy and Physiology - Kidney - ForKristelle ModalesÎncă nu există evaluări
- Renal MCQ 1Document2 paginiRenal MCQ 1AzizÎncă nu există evaluări
- Disorders of Sodium and Potassium MetabolismDocument18 paginiDisorders of Sodium and Potassium MetabolismTukuWoÎncă nu există evaluări
- Urinary EliminationDocument7 paginiUrinary EliminationMarcus, RN100% (14)
- Class - X Sample Paper PDFDocument16 paginiClass - X Sample Paper PDFApex Institute0% (1)
- 300 FCPS MCQS Solved 2Document14 pagini300 FCPS MCQS Solved 2AS IFÎncă nu există evaluări
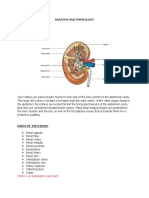
- The Kidneys Are A Pair of Purplish-Brown Organs Is Located Below The Ribs Toward The Middle ofDocument6 paginiThe Kidneys Are A Pair of Purplish-Brown Organs Is Located Below The Ribs Toward The Middle ofJoana Bless PereyÎncă nu există evaluări
- Urinary System (2015 - 06 - 09 22 - 21 - 55 Utc)Document44 paginiUrinary System (2015 - 06 - 09 22 - 21 - 55 Utc)DiazÎncă nu există evaluări
- Class 11th Human Physio TestDocument8 paginiClass 11th Human Physio TestSaurabh KrishnaÎncă nu există evaluări
- Nursing Case Study (Final) - ArchieDocument23 paginiNursing Case Study (Final) - ArchieArchie Astorga100% (2)
- Thesis On Diet and NutritionDocument7 paginiThesis On Diet and NutritionScott Bou100% (1)
- Excretion: Excretion. Elimination of Drugs and Its Metabolites FromDocument18 paginiExcretion: Excretion. Elimination of Drugs and Its Metabolites FromAlphahin 17100% (1)
- Life Sciences Grade 11 Revision Terms 3 and 4 2021Document42 paginiLife Sciences Grade 11 Revision Terms 3 and 4 2021Letlhogonolo RaotsileÎncă nu există evaluări
- Excretion and Osmoregulation in FishesDocument15 paginiExcretion and Osmoregulation in FishesMubashar Hussain100% (2)
- Mechanisms of Glomerular Crescent FormationDocument19 paginiMechanisms of Glomerular Crescent Formation_siegfried_Încă nu există evaluări
- Biokimia Urine, Sisa Metabolisme Dan Pembentukan BatuDocument104 paginiBiokimia Urine, Sisa Metabolisme Dan Pembentukan BatuHenny Taufik GosalÎncă nu există evaluări
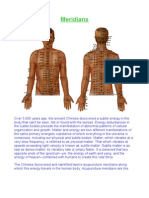
- MeridiansDocument11 paginiMeridiansa2017399883% (6)
- Edited - The Excretory System WorksheetDocument4 paginiEdited - The Excretory System WorksheethmalmashaanÎncă nu există evaluări
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedDe la EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedEvaluare: 5 din 5 stele5/5 (80)
- The Obesity Code: Unlocking the Secrets of Weight LossDe la EverandThe Obesity Code: Unlocking the Secrets of Weight LossEvaluare: 4 din 5 stele4/5 (6)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDDe la EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDEvaluare: 5 din 5 stele5/5 (1)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeDe la EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeEvaluare: 2 din 5 stele2/5 (1)
- The Age of Magical Overthinking: Notes on Modern IrrationalityDe la EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityEvaluare: 4 din 5 stele4/5 (24)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionDe la EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionEvaluare: 4 din 5 stele4/5 (404)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisDe la EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisEvaluare: 4.5 din 5 stele4.5/5 (42)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsDe la EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsEvaluare: 5 din 5 stele5/5 (1)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaDe la EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaEvaluare: 4.5 din 5 stele4.5/5 (266)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisDe la EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisEvaluare: 4 din 5 stele4/5 (1)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsDe la EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsÎncă nu există evaluări
- Why We Die: The New Science of Aging and the Quest for ImmortalityDe la EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityEvaluare: 4 din 5 stele4/5 (3)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryDe la EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryEvaluare: 4 din 5 stele4/5 (44)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.De la EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Evaluare: 4.5 din 5 stele4.5/5 (110)
- Gut: the new and revised Sunday Times bestsellerDe la EverandGut: the new and revised Sunday Times bestsellerEvaluare: 4 din 5 stele4/5 (392)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDe la EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisEvaluare: 3.5 din 5 stele3.5/5 (2)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsDe la EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsEvaluare: 3.5 din 5 stele3.5/5 (3)
- The Marshmallow Test: Mastering Self-ControlDe la EverandThe Marshmallow Test: Mastering Self-ControlEvaluare: 4.5 din 5 stele4.5/5 (58)
- Dark Psychology: Learn To Influence Anyone Using Mind Control, Manipulation And Deception With Secret Techniques Of Dark Persuasion, Undetected Mind Control, Mind Games, Hypnotism And BrainwashingDe la EverandDark Psychology: Learn To Influence Anyone Using Mind Control, Manipulation And Deception With Secret Techniques Of Dark Persuasion, Undetected Mind Control, Mind Games, Hypnotism And BrainwashingEvaluare: 4 din 5 stele4/5 (1138)
- Sleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningDe la EverandSleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningEvaluare: 4 din 5 stele4/5 (3)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsDe la EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsEvaluare: 4.5 din 5 stele4.5/5 (169)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessDe la EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessEvaluare: 4.5 din 5 stele4.5/5 (328)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDe la EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsEvaluare: 4.5 din 5 stele4.5/5 (6)