S-ar putea să vă placă și
- The Biology of Cancer: Second EditionDocument47 paginiThe Biology of Cancer: Second EditionxbÎncă nu există evaluări
- CAPE PhotosynthesisDocument20 paginiCAPE PhotosynthesisIsheba Warren100% (24)
- Ethem Murat Arsava (Eds.) - Nutrition in Neurologic Disorders - A Practical Guide (2017, Springer International Publishing)Document209 paginiEthem Murat Arsava (Eds.) - Nutrition in Neurologic Disorders - A Practical Guide (2017, Springer International Publishing)riddhiÎncă nu există evaluări
- Myoglobin & Hemoglobin: Essential Oxygen Transport ProteinsDocument53 paginiMyoglobin & Hemoglobin: Essential Oxygen Transport ProteinsAgnetasia PriscilliaÎncă nu există evaluări
- Chloroplasts Structure and Function FactsheetDocument4 paginiChloroplasts Structure and Function Factsheettapas kundu100% (1)
- Compare Mitosis and Meiosi1Document9 paginiCompare Mitosis and Meiosi1Reuben John AbalayanÎncă nu există evaluări
- Human Anatomy 9th Edition Martini Test BankDocument31 paginiHuman Anatomy 9th Edition Martini Test BankKevinHarrisoncatjn100% (10)
- Photosynthesis: How Plants Make FoodDocument9 paginiPhotosynthesis: How Plants Make FoodJulia Clarisse Alanza BallonÎncă nu există evaluări
- SCI 8 Q4 MOD 2 Cellular Reproduction NKADocument21 paginiSCI 8 Q4 MOD 2 Cellular Reproduction NKAhv jjÎncă nu există evaluări
- Onion Root Mitosis LabDocument6 paginiOnion Root Mitosis Labapi-2469736100% (1)
- Phloem: - Sieve Tube Elements (Angiosperms) - Sieve Cells (Gymnosperms)Document9 paginiPhloem: - Sieve Tube Elements (Angiosperms) - Sieve Cells (Gymnosperms)xbÎncă nu există evaluări
- Stomatal BiologyDocument11 paginiStomatal BiologyxbÎncă nu există evaluări
- 1.2.3 Transport in PlantsDocument11 pagini1.2.3 Transport in PlantsMo_Bash1Încă nu există evaluări
- A Level Transport FinestDocument25 paginiA Level Transport FinestBaguma alexÎncă nu există evaluări
- Mass Transport PlantsDocument63 paginiMass Transport Plantssharanya.10.puneet.8.karolÎncă nu există evaluări
- Photosynthesis - Part 1Document90 paginiPhotosynthesis - Part 1Antonio WilloughbyÎncă nu există evaluări
- μm) w high fidelity, optically transparent to ~300 nm,: l = (vep + veo) ti = (ep + eo) Viti / LDocument3 paginiμm) w high fidelity, optically transparent to ~300 nm,: l = (vep + veo) ti = (ep + eo) Viti / LImma WongÎncă nu există evaluări
- EEB 162 Final Study GuideDocument13 paginiEEB 162 Final Study GuideSydney ChangÎncă nu există evaluări
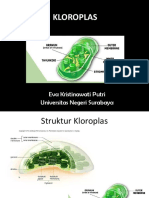
- KLOROPLASDocument32 paginiKLOROPLASSintha Eka AshariÎncă nu există evaluări
- Research Paper On Plant BiochemistryDocument8 paginiResearch Paper On Plant Biochemistrygz9g97ha100% (1)
- Photosynthesis, Respiration, and Long-Distance TransportDocument2 paginiPhotosynthesis, Respiration, and Long-Distance TransportAlv1en HidayatÎncă nu există evaluări
- Bio2 90464 Cell KeywordsDocument6 paginiBio2 90464 Cell Keywordsapi-277699810Încă nu există evaluări
- Flow cytometry analysis and cell sorting in plants: past developments and future potentialDocument6 paginiFlow cytometry analysis and cell sorting in plants: past developments and future potentialTallie ZeidlerÎncă nu există evaluări
- Structure of A ChloroplastDocument2 paginiStructure of A ChloroplastsreenitthiÎncă nu există evaluări
- Biology Unit 4 Revision: Chloroplast Structure and FunctionDocument2 paginiBiology Unit 4 Revision: Chloroplast Structure and FunctionsreenitthiÎncă nu există evaluări
- Chloroplast Structure and FunctionDocument19 paginiChloroplast Structure and FunctionFizul HelmiÎncă nu există evaluări
- Allium Cepa (Onion) Capsicum (Pepper) Hydrilla Sprig Solanum Lycopersicum (Tomato) Cucurbita (Squash) ShootDocument3 paginiAllium Cepa (Onion) Capsicum (Pepper) Hydrilla Sprig Solanum Lycopersicum (Tomato) Cucurbita (Squash) ShootMEDELYN KEITH ESTANISLAOÎncă nu există evaluări
- Functions of The Cell MembraneDocument5 paginiFunctions of The Cell Membranemekusa10Încă nu există evaluări
- 2014 Nature and Composition of Plant Ag Review 2013Document250 pagini2014 Nature and Composition of Plant Ag Review 2013Mc Wilson DecenaÎncă nu există evaluări
- Chloroplast Structure and FunctionDocument31 paginiChloroplast Structure and FunctionShikhaj ArmaanÎncă nu există evaluări
- Chloroplast and Photosynthesis: For Advanced Cell and Molecular BiologyDocument33 paginiChloroplast and Photosynthesis: For Advanced Cell and Molecular BiologyJean Rose GenovaÎncă nu există evaluări
- PhotosynthesisDocument35 paginiPhotosynthesisYuh moddaÎncă nu există evaluări
- Overview: Cellular Messaging: © 2011 Pearson Education, IncDocument69 paginiOverview: Cellular Messaging: © 2011 Pearson Education, IncbbyiknowÎncă nu există evaluări
- K12 STEM Training on Cell TransportDocument53 paginiK12 STEM Training on Cell Transportpotato macchiatoÎncă nu există evaluări
- Lecture-1 Translocation of FoodDocument31 paginiLecture-1 Translocation of FoodMuhammad Arshad Ali BhuttaÎncă nu există evaluări
- Chloroplast 1.2Document5 paginiChloroplast 1.2Ishita Kumari100% (1)
- A M I E: Dvanced Ethods N LectrophysiologyDocument43 paginiA M I E: Dvanced Ethods N LectrophysiologyArtur LiberatoÎncă nu există evaluări
- Unit 4 Notes BioDocument130 paginiUnit 4 Notes Biohari chohanÎncă nu există evaluări
- Histology: Cytology, The Study of The Structure and Function of CellsDocument48 paginiHistology: Cytology, The Study of The Structure and Function of CellsZmeu CristinaÎncă nu există evaluări
- Onion root cell mitosis effects of acetoorcein staining durationDocument4 paginiOnion root cell mitosis effects of acetoorcein staining durationFakh IraÎncă nu există evaluări
- CELLS: Structure and Function of the Building Blocks of LifeDocument5 paginiCELLS: Structure and Function of the Building Blocks of LifegraceÎncă nu există evaluări
- Transport in PlantsDocument18 paginiTransport in PlantsMostafa A MansiÎncă nu există evaluări
- scsc7211's Version of Alan's DAT Biology NotesDocument47 paginiscsc7211's Version of Alan's DAT Biology NotesMusawir YasinÎncă nu există evaluări
- CHLOROPLASTDocument4 paginiCHLOROPLASTjyotirmoy choudhuryÎncă nu există evaluări
- Lec. 01.introductionDocument43 paginiLec. 01.introduction6ng2q6xywwÎncă nu există evaluări
- Plants use photosynthesis products for growth and fuelDocument6 paginiPlants use photosynthesis products for growth and fuelJanneHillaryÎncă nu există evaluări
- Photosynthesis OFFICIALDocument36 paginiPhotosynthesis OFFICIALAsawni McDowellÎncă nu există evaluări
- CELL PHYSIOLOGY@starDocument6 paginiCELL PHYSIOLOGY@starDENIS KIPKOSGEIÎncă nu există evaluări
- Lecture 1Document20 paginiLecture 1ALL ABOUT PAGEANTARYÎncă nu există evaluări
- CRSC 1 GUIDE QUESTIONS FOR CHAPTER 4Document5 paginiCRSC 1 GUIDE QUESTIONS FOR CHAPTER 4RanielJohn CollantesÎncă nu există evaluări
- Def PhysioDocument34 paginiDef PhysioFrancis Ben BerjaÎncă nu există evaluări
- Pathogen Vs Cell MembraneDocument12 paginiPathogen Vs Cell MembraneHunAin SajjAdÎncă nu există evaluări
- ANPH-M1-CU2. The Cell & Cellular Metabolism and Reproduction (Mitosis and Meiosis) PDFDocument26 paginiANPH-M1-CU2. The Cell & Cellular Metabolism and Reproduction (Mitosis and Meiosis) PDFJamaica M DanguecanÎncă nu există evaluări
- COGS 101A: Sensation and Perception: Virginia R. de Sa Department of Cognitive Science Ucsd The Retina - Chapter 2Document39 paginiCOGS 101A: Sensation and Perception: Virginia R. de Sa Department of Cognitive Science Ucsd The Retina - Chapter 2Ratnaganadi ParamitaÎncă nu există evaluări
- Plant Cell SignalingDocument35 paginiPlant Cell SignalingAmelia Sukma Wati325Încă nu există evaluări
- Photosynthesis and ATP Synthesis Part 1Document39 paginiPhotosynthesis and ATP Synthesis Part 1Andy100% (1)
- Breanna 2Document2 paginiBreanna 2AbhishekTyagiÎncă nu există evaluări
- Plant Physiology EvaluationsDocument13 paginiPlant Physiology Evaluationslisanames.23Încă nu există evaluări
- Topic 1.1 - Cell TheoryDocument6 paginiTopic 1.1 - Cell Theoryjahajaha_svensson609Încă nu există evaluări
- Lection 1 and 2Document2 paginiLection 1 and 2MariaÎncă nu există evaluări
- The Cell and Cellular Metabolism ReproductionDocument4 paginiThe Cell and Cellular Metabolism ReproductionApple AbriamÎncă nu există evaluări
- BIOL111 1st Sem, Final TermDocument40 paginiBIOL111 1st Sem, Final TermJasmine CatanaÎncă nu există evaluări
- Phloem: Molecular Cell Biology, Systemic Communication, Biotic InteractionsDe la EverandPhloem: Molecular Cell Biology, Systemic Communication, Biotic InteractionsÎncă nu există evaluări
- The Journal of PathologyDocument14 paginiThe Journal of PathologyxbÎncă nu există evaluări
- Review: RAS Proteins and Their Regulators in Human DiseaseDocument17 paginiReview: RAS Proteins and Their Regulators in Human DiseasexbÎncă nu există evaluări
- Protein Modification Second Messengers: Signal Transduction May Be Cell Autonomous or Non-Cell AutonomousDocument16 paginiProtein Modification Second Messengers: Signal Transduction May Be Cell Autonomous or Non-Cell AutonomousxbÎncă nu există evaluări
- Seeds, Seedlings, and Early DevelopmentDocument45 paginiSeeds, Seedlings, and Early DevelopmentxbÎncă nu există evaluări
- Plant Cell Walls - Complex Structures Involved In: Repens)Document21 paginiPlant Cell Walls - Complex Structures Involved In: Repens)xbÎncă nu există evaluări
- Assimilation of Mineral Nutrients in PlantsDocument11 paginiAssimilation of Mineral Nutrients in PlantsxbÎncă nu există evaluări
- Reactions of Plant Glycolysis and FermentationDocument8 paginiReactions of Plant Glycolysis and FermentationxbÎncă nu există evaluări
- Mind Maps in Biochemistry - (Chemistry of Proteins)Document6 paginiMind Maps in Biochemistry - (Chemistry of Proteins)Gus LionsÎncă nu există evaluări
- CorneaDocument41 paginiCorneaNikhil KorripatiÎncă nu există evaluări
- Prokaryotic CellDocument2 paginiProkaryotic CellFrancine Dominique CollantesÎncă nu există evaluări
- Methods: Aurélia Battesti, Emmanuelle BouveretDocument10 paginiMethods: Aurélia Battesti, Emmanuelle BouveretrezqÎncă nu există evaluări
- VanPutte Seeleys Essentials 11e Chap07 PPT AccessibleDocument140 paginiVanPutte Seeleys Essentials 11e Chap07 PPT AccessibleCyrille Aira AndresaÎncă nu există evaluări
- Cell Lysis ProtocolDocument5 paginiCell Lysis ProtocolmnstnÎncă nu există evaluări
- Laboratory Activity #1 The Cell (1) (1) 2Document3 paginiLaboratory Activity #1 The Cell (1) (1) 2Kim VenturaÎncă nu există evaluări
- Mummy SyllabusDocument89 paginiMummy SyllabusHarshavardhan BGÎncă nu există evaluări
- IGCSE Student Revision Power Point Topic 5 - 複本Document16 paginiIGCSE Student Revision Power Point Topic 5 - 複本yt kÎncă nu există evaluări
- 2 Types of Media Culture in Animal Cell: Natural and Artificial MediaDocument11 pagini2 Types of Media Culture in Animal Cell: Natural and Artificial MediaSandeep TanwarÎncă nu există evaluări
- Sbi3C Name: Characteristics of Bacteria Worksheet: Big Idea! IdeasDocument2 paginiSbi3C Name: Characteristics of Bacteria Worksheet: Big Idea! IdeasPaulina JakubiecÎncă nu există evaluări
- E) Two: Shah Kamal Institute Thatta (Skit)Document2 paginiE) Two: Shah Kamal Institute Thatta (Skit)Wahaj KhanÎncă nu există evaluări
- Southwestern University Protein Structure QuizDocument4 paginiSouthwestern University Protein Structure QuizAndrew DonaireÎncă nu există evaluări
- Lesson 4 - Non-Membrane-bound OrganellesDocument3 paginiLesson 4 - Non-Membrane-bound OrganellesEarl Caesar Quiba PagunsanÎncă nu există evaluări
- Connective Tissue NotesDocument6 paginiConnective Tissue Notesferdinand padillaÎncă nu există evaluări
- ML Undergraduate Research Symposium Presentation - Madison LuceDocument1 paginăML Undergraduate Research Symposium Presentation - Madison LucenurjÎncă nu există evaluări
- Chapter 6 A Tour of The CellDocument3 paginiChapter 6 A Tour of The CellMohammad Mahrus Bin AshfaqÎncă nu există evaluări
- Quiz #1 Questions A&P 1Document14 paginiQuiz #1 Questions A&P 1trick49Încă nu există evaluări
- 2012 Smarcal1 Human Molecular GeneticsDocument867 pagini2012 Smarcal1 Human Molecular Geneticsrameshramyb100% (1)
- DNA Polymerase NotesDocument10 paginiDNA Polymerase NoteslordniklausÎncă nu există evaluări
- Cell anatomy and divisionDocument4 paginiCell anatomy and divisionMarielle SaymanÎncă nu există evaluări
- AgNPs Paper 3Document9 paginiAgNPs Paper 3VILEOLAGOLDÎncă nu există evaluări
- Parts of the nerve cell and their functionsDocument5 paginiParts of the nerve cell and their functionsAaron WallaceÎncă nu există evaluări
- Meiosis: How Gametes Form with Half the ChromosomesDocument47 paginiMeiosis: How Gametes Form with Half the ChromosomesRhaine EstebanÎncă nu există evaluări
- RP - MicrosDocument17 paginiRP - MicrosNeel MutagiÎncă nu există evaluări