S-ar putea să vă placă și
- Salicylic Acid and Photosynthesis: Signalling and EffectsDocument31 paginiSalicylic Acid and Photosynthesis: Signalling and EffectsKanishk KannaÎncă nu există evaluări
- 12856-Article Text-45865-1-10-20190909Document12 pagini12856-Article Text-45865-1-10-20190909Gutierrez DieguiittoÎncă nu există evaluări
- Salicylic Acid Signalling in Plants: Molecular SciencesDocument6 paginiSalicylic Acid Signalling in Plants: Molecular SciencesKanishk KannaÎncă nu există evaluări
- Auxin PDFDocument16 paginiAuxin PDFvimalrajÎncă nu există evaluări
- Sulfur 2019Document5 paginiSulfur 2019Prajna AdityaraniÎncă nu există evaluări
- The Biological Action of Saponins in Animal SystemsDocument19 paginiThe Biological Action of Saponins in Animal SystemsAlejandro Rivera Guzmán100% (1)
- Plants 11 01776Document16 paginiPlants 11 01776bela OktarindaÎncă nu există evaluări
- Regulation of Water, Salinity, and Cold Stress Responses by Salicylic AcidDocument12 paginiRegulation of Water, Salinity, and Cold Stress Responses by Salicylic AcidBastian SeguelÎncă nu există evaluări
- Silvaetal 2017selenium1Document11 paginiSilvaetal 2017selenium1Florin SJÎncă nu există evaluări
- Plant SaponinsDocument25 paginiPlant SaponinsCamila ScoparoÎncă nu există evaluări
- 1 s2.0 S1878535220302707 MainDocument9 pagini1 s2.0 S1878535220302707 Mainaryestukan03Încă nu există evaluări
- Plant Hormones and Nutrient Signaling: Ó Springer Science+Business Media B.V. 2008Document13 paginiPlant Hormones and Nutrient Signaling: Ó Springer Science+Business Media B.V. 2008Isabella Perez MendezÎncă nu există evaluări
- 10 1002@cbdv 200800070Document254 pagini10 1002@cbdv 200800070Duong Pham QuangÎncă nu există evaluări
- Antioxidants 11 01828Document14 paginiAntioxidants 11 01828J MrÎncă nu există evaluări
- 3 KFX ZXR PPNK 9 H K84 ZKGFBCNDocument10 pagini3 KFX ZXR PPNK 9 H K84 ZKGFBCNAlisson CirinoÎncă nu există evaluări
- Agronomy 11 00500 v3Document18 paginiAgronomy 11 00500 v3Lily LiÎncă nu există evaluări
- Final ThesisDocument51 paginiFinal ThesisUtkarsh bajpaiÎncă nu există evaluări
- Role of Salicylic Acid (SA) in Plants - A Review: S Muthulakshmi and K LingakumarDocument5 paginiRole of Salicylic Acid (SA) in Plants - A Review: S Muthulakshmi and K LingakumarEinstein Albert0% (1)
- Impact of Saponins or Saponin-Containing Plant Materials On Ruminant Production, A Review - Wina (Document13 paginiImpact of Saponins or Saponin-Containing Plant Materials On Ruminant Production, A Review - Wina (Sylvain KerrosÎncă nu există evaluări
- Emerging Plant Growth Regulators in Agriculture: Roles in Stress ToleranceDe la EverandEmerging Plant Growth Regulators in Agriculture: Roles in Stress ToleranceM. NaeemÎncă nu există evaluări
- Jasmonic Acid Signaling Pathway in Response To Abiotic Stresses in PlantsDocument19 paginiJasmonic Acid Signaling Pathway in Response To Abiotic Stresses in PlantsrachiiidaÎncă nu există evaluări
- Integrated Physiological, Metabolite and Proteomic Analysis Reveal TheDocument14 paginiIntegrated Physiological, Metabolite and Proteomic Analysis Reveal TheHENNIO GILBERTO TRUJILLO RAMONÎncă nu există evaluări
- Antioxidant and Cytotoxic Activities of Centella Asiatica (L) UrbDocument9 paginiAntioxidant and Cytotoxic Activities of Centella Asiatica (L) UrbUwais AlÎncă nu există evaluări
- Structures of Astaxanthin and Their Consequences For Therapeutic ApplicationDocument16 paginiStructures of Astaxanthin and Their Consequences For Therapeutic ApplicationHelenaÎncă nu există evaluări
- Captura de Pantalla 2023-12-10 A La(s) 13.37.26Document17 paginiCaptura de Pantalla 2023-12-10 A La(s) 13.37.26Ronald FarinangoÎncă nu există evaluări
- Genes and Enzymes of Carotenoid Biosynthesis in PlantsDocument14 paginiGenes and Enzymes of Carotenoid Biosynthesis in PlantsskljoleÎncă nu există evaluări
- Salt Tolerance in Plants-Role of Osmolytes and Osmoprotectant-Vijayata Singh and Jogendra SinghDocument5 paginiSalt Tolerance in Plants-Role of Osmolytes and Osmoprotectant-Vijayata Singh and Jogendra SinghDr. Vijayata Singh0% (1)
- Acacia Mangium Willd (Acácia) Extract Antioxidant and Antiproliferative ActivitiesDocument10 paginiAcacia Mangium Willd (Acácia) Extract Antioxidant and Antiproliferative ActivitiesSabrina JonesÎncă nu există evaluări
- Thesis On Antioxidant Activity of Medicinal Plants PDFDocument6 paginiThesis On Antioxidant Activity of Medicinal Plants PDFafazapfjl100% (2)
- Functions of Jasmonic Acid in Plant Regulation and Response To Abiotic StressDocument17 paginiFunctions of Jasmonic Acid in Plant Regulation and Response To Abiotic Stressutama 3002Încă nu există evaluări
- Molecules 26 00174 v2Document28 paginiMolecules 26 00174 v2Daniel HernándezÎncă nu există evaluări
- Variability of Hordeum Vulgare L. Cultivars in Yield, Antioxidant Potential, and Cholinesterase Inhibitory ActivityDocument14 paginiVariability of Hordeum Vulgare L. Cultivars in Yield, Antioxidant Potential, and Cholinesterase Inhibitory Activitydhafer alhaidaryÎncă nu există evaluări
- Molecules: Lamium Plants-A Comprehensive Review On Health Benefits and Biological ActivitiesDocument23 paginiMolecules: Lamium Plants-A Comprehensive Review On Health Benefits and Biological ActivitiesAd NeagoieÎncă nu există evaluări
- Jurnal SPT 2Document8 paginiJurnal SPT 2Alma SafiraÎncă nu există evaluări
- Saur Inhibition of Pp2C-D Phosphatases Activates Plasma Membrane H - Atpases To Promote Cell Expansion in ArabidopsisDocument14 paginiSaur Inhibition of Pp2C-D Phosphatases Activates Plasma Membrane H - Atpases To Promote Cell Expansion in ArabidopsisjustfortrashjunkÎncă nu există evaluări
- BalachandaranDocument10 paginiBalachandaranDrGomathi Sankar KandappanÎncă nu există evaluări
- Molecules 26 05347 v2Document16 paginiMolecules 26 05347 v2LuisEduardoRomeroÎncă nu există evaluări
- Content ServerDocument15 paginiContent ServerZeyar Than WinÎncă nu există evaluări
- Annual Plant Reviews, Phosphorus Metabolism in PlantsDe la EverandAnnual Plant Reviews, Phosphorus Metabolism in PlantsWilliam PlaxtonÎncă nu există evaluări
- Antioxidants 10 01876 v2Document37 paginiAntioxidants 10 01876 v2VILEOLAGOLDÎncă nu există evaluări
- Article STDocument17 paginiArticle STHanish JainÎncă nu există evaluări
- Metabolism and Interactions: The Chemistry and Biology of Compounds Containing Amino SugarsDe la EverandMetabolism and Interactions: The Chemistry and Biology of Compounds Containing Amino SugarsEndre A. BalazsÎncă nu există evaluări
- Antioxidant - Activity - of Pereskia BleoDocument6 paginiAntioxidant - Activity - of Pereskia BleoAvely AntoniusÎncă nu există evaluări
- The Biological Action of Saponins in Animal Systems A ReviewDocument19 paginiThe Biological Action of Saponins in Animal Systems A ReviewCorinaÎncă nu există evaluări
- Isolation and Antioxidant Activity of Flavonoids From HolarrhenaDocument6 paginiIsolation and Antioxidant Activity of Flavonoids From HolarrhenaIra MonicaÎncă nu există evaluări
- Review Jurnal Faktor LingkunganDocument26 paginiReview Jurnal Faktor LingkunganyunitaknÎncă nu există evaluări
- Pyro X A Sulfone MetabolismDocument6 paginiPyro X A Sulfone MetabolismJuan Pablo EspinosaÎncă nu există evaluări
- Structural and Biological Diversity of Cyclic Octadecanoids, Jasmonates, and MimeticsDocument2 paginiStructural and Biological Diversity of Cyclic Octadecanoids, Jasmonates, and MimeticsFranklin Edmundo Quenguan CuaranÎncă nu există evaluări
- Total Reactive Antioxidant Potential (TRAP) and Total Antioxidant Reactivity (TAR) of Medicinal Plants Used in Southwest Amazonia (Bolivia and Peru)Document10 paginiTotal Reactive Antioxidant Potential (TRAP) and Total Antioxidant Reactivity (TAR) of Medicinal Plants Used in Southwest Amazonia (Bolivia and Peru)Betty Isabel Blanco CamposÎncă nu există evaluări
- Silicon-Mediated Accumulation of Flavonoid Phytoalexins in CucumberDocument6 paginiSilicon-Mediated Accumulation of Flavonoid Phytoalexins in CucumberRichard Quispe QuispeÎncă nu există evaluări
- Pyrethroids: Mammalian Metabolism and Toxicity: Hideo KanekoDocument6 paginiPyrethroids: Mammalian Metabolism and Toxicity: Hideo KanekoNacho BressánÎncă nu există evaluări
- Ajps - 2014112818074195 (2014)Document11 paginiAjps - 2014112818074195 (2014)hrishikesh08Încă nu există evaluări
- Auxin Production As An Integrator of Environmental Cues For Developmental Growth RegulationDocument12 paginiAuxin Production As An Integrator of Environmental Cues For Developmental Growth RegulationSylviaYanaÎncă nu există evaluări
- Govt. College University, Faisalabad Department of Botany Synopsis For M. Phil. DegreeDocument14 paginiGovt. College University, Faisalabad Department of Botany Synopsis For M. Phil. DegreehabibfatimaÎncă nu există evaluări
- 2016 Art96Document23 pagini2016 Art96Nguyễn Ngọc NamÎncă nu există evaluări
- Comparative Free Radical Scavenging Potentials of Different Parts ofDocument6 paginiComparative Free Radical Scavenging Potentials of Different Parts ofTito saragihÎncă nu există evaluări
- Comparative Free Radical Scavenging Potentials of Different Parts ofDocument6 paginiComparative Free Radical Scavenging Potentials of Different Parts ofTito saragihÎncă nu există evaluări
- tmp6198 TMPDocument14 paginitmp6198 TMPFrontiersÎncă nu există evaluări
- Agri Paper 4Document12 paginiAgri Paper 4VikasÎncă nu există evaluări
- E3sconf - Ichm13 - 13005 AmaranthusDocument4 paginiE3sconf - Ichm13 - 13005 AmaranthusrachiiidaÎncă nu există evaluări
- Chemosphere: María T. Domínguez, Paula Madej On, Engracia Madej On, Manuel J. DiazDocument9 paginiChemosphere: María T. Domínguez, Paula Madej On, Engracia Madej On, Manuel J. DiazrachiiidaÎncă nu există evaluări
- 167 - Phytoremediation Cistus, Dittrichia, Euphorbia (2011)Document7 pagini167 - Phytoremediation Cistus, Dittrichia, Euphorbia (2011)rachiiidaÎncă nu există evaluări
- Heavy Metals Concentrations and Risk Assessment of Rose - 2017 - Annals of AgricDocument6 paginiHeavy Metals Concentrations and Risk Assessment of Rose - 2017 - Annals of AgricrachiiidaÎncă nu există evaluări
- Phytoremediation Techniques in Wastewater Treatment: Milena Materac, Anna Wyrwicka, El Bieta SobieckaDocument10 paginiPhytoremediation Techniques in Wastewater Treatment: Milena Materac, Anna Wyrwicka, El Bieta SobieckarachiiidaÎncă nu există evaluări
- 2011 Ijms-12-07760 Brassica in Phytoremediation and BiofumigationDocument12 pagini2011 Ijms-12-07760 Brassica in Phytoremediation and BiofumigationrachiiidaÎncă nu există evaluări
- Comparative Assessment For Hyperaccumulatory and Phytoremediation Capability of Three Wild WeedsDocument11 paginiComparative Assessment For Hyperaccumulatory and Phytoremediation Capability of Three Wild WeedsrachiiidaÎncă nu există evaluări
- Plant Physiology and Biochemistry: ThalianaDocument8 paginiPlant Physiology and Biochemistry: ThalianarachiiidaÎncă nu există evaluări
- Mutation Research/Genetic Toxicology and Environmental MutagenesisDocument7 paginiMutation Research/Genetic Toxicology and Environmental MutagenesisrachiiidaÎncă nu există evaluări
- Germination Rate of Stinkwort (Dittrichia Graveolens) and False Yellowhead (D. Viscosa) in Relation To SalinityDocument8 paginiGermination Rate of Stinkwort (Dittrichia Graveolens) and False Yellowhead (D. Viscosa) in Relation To SalinityrachiiidaÎncă nu există evaluări
- Notes On The Phenology of Dittrichia ViscosaDocument10 paginiNotes On The Phenology of Dittrichia ViscosarachiiidaÎncă nu există evaluări
- Contribution of The Arbuscular Mycorrhizal Symbiosis To Heavy Metal PhytoremediationDocument8 paginiContribution of The Arbuscular Mycorrhizal Symbiosis To Heavy Metal PhytoremediationrachiiidaÎncă nu există evaluări
- Plant Life History and Above-Belowground Interactions: Missing LinksDocument11 paginiPlant Life History and Above-Belowground Interactions: Missing LinksrachiiidaÎncă nu există evaluări
- Karel Et Al 2019Document19 paginiKarel Et Al 2019rachiiidaÎncă nu există evaluări
- Silva Et Al 2011Document13 paginiSilva Et Al 2011rachiiidaÎncă nu există evaluări
- Saini2014 Article ElicitorsSAAndMJEnhanceCarotenDocument10 paginiSaini2014 Article ElicitorsSAAndMJEnhanceCarotenrachiiidaÎncă nu există evaluări
- BALLAR - 2009-Plant, Cell & EnvironmentDocument13 paginiBALLAR - 2009-Plant, Cell & EnvironmentrachiiidaÎncă nu există evaluări
- Environmental and Experimental BotanyDocument8 paginiEnvironmental and Experimental BotanyrachiiidaÎncă nu există evaluări
- MCP 049Document11 paginiMCP 049rachiiidaÎncă nu există evaluări
- The Endophytic System of Mediterranean Cytinus (Cytinaceae) Developing On Five Host Cistaceae SpeciesDocument9 paginiThe Endophytic System of Mediterranean Cytinus (Cytinaceae) Developing On Five Host Cistaceae SpeciesrachiiidaÎncă nu există evaluări
- Holoparasite Heavymetals CistusDocument9 paginiHoloparasite Heavymetals CistusrachiiidaÎncă nu există evaluări
- Women and Islam Fatima Mernisi PDFDocument238 paginiWomen and Islam Fatima Mernisi PDFrachiiidaÎncă nu există evaluări
- Jasmonic Acid Signaling Pathway in Response To Abiotic Stresses in PlantsDocument19 paginiJasmonic Acid Signaling Pathway in Response To Abiotic Stresses in PlantsrachiiidaÎncă nu există evaluări
- The Ant-Pollination System of Cytinus Hypocistis (Cytinaceae), A Mediterranean Root HoloparasiteDocument11 paginiThe Ant-Pollination System of Cytinus Hypocistis (Cytinaceae), A Mediterranean Root HoloparasiterachiiidaÎncă nu există evaluări
- Platinum Formulary 2016Document61 paginiPlatinum Formulary 2016jÎncă nu există evaluări
- AromaticityDocument12 paginiAromaticityV G Viju KumarÎncă nu există evaluări
- Organic Molecule:: Nucleic AcidsDocument26 paginiOrganic Molecule:: Nucleic AcidsCharles SalcedoÎncă nu există evaluări
- A2 4 1 1 Krebs CycleDocument12 paginiA2 4 1 1 Krebs CycleHo Man ChanÎncă nu există evaluări
- CHEM 2324 SPRING 2012 EXAM 1 W AnswersDocument8 paginiCHEM 2324 SPRING 2012 EXAM 1 W AnswersSusana PortilloÎncă nu există evaluări
- Siklus KalvinDocument5 paginiSiklus Kalvinnovani florensiaÎncă nu există evaluări
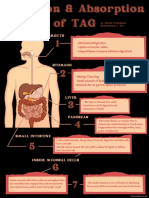
- Digestion & Absorption of TAGDocument1 paginăDigestion & Absorption of TAGJanine Franchesca SuministradoÎncă nu există evaluări
- Science Driven NutritionDocument39 paginiScience Driven Nutritionjan100% (1)
- TB Chapter18Document20 paginiTB Chapter18Ruchi Briam James LagitnayÎncă nu există evaluări
- Carbohydrates 2Document81 paginiCarbohydrates 2smcm11Încă nu există evaluări
- Bch3701 Answer Sheet CompleteDocument14 paginiBch3701 Answer Sheet CompleteNdumiso NdawondeÎncă nu există evaluări
- Lecture 1 by Madam Shagufta PolymerDocument32 paginiLecture 1 by Madam Shagufta PolymerAhmad Shah 8999Încă nu există evaluări
- Protein Synthesis Drag & Drop Activity 2021Document6 paginiProtein Synthesis Drag & Drop Activity 2021Gloria LaneÎncă nu există evaluări
- Dong Dong New Product ListDocument3 paginiDong Dong New Product ListPriyank PataniÎncă nu există evaluări
- GluconeogenesisDocument14 paginiGluconeogenesisapi-453481795Încă nu există evaluări
- Urea CycleDocument13 paginiUrea CycleShampa SenÎncă nu există evaluări
- Protein Lesson PlanDocument4 paginiProtein Lesson Planapi-270183943Încă nu există evaluări
- +++ Fundamentals of Petroleum RefiningDocument6 pagini+++ Fundamentals of Petroleum Refiningnadjib62Încă nu există evaluări
- Fat MetabolismDocument14 paginiFat MetabolismDimakatso Nkanyezi DubazaneÎncă nu există evaluări
- Pediatric Practice Math Problems Answer KeyDocument3 paginiPediatric Practice Math Problems Answer KeyKaren Hutchinson50% (2)
- Triatmo Budiyuwono SPJP (K), FIHA, FAPSICDocument40 paginiTriatmo Budiyuwono SPJP (K), FIHA, FAPSICganangahimsaÎncă nu există evaluări
- Food and Drug Administration, HHS - Food & Beverage FDA 21 C.F.R. 175.300Document18 paginiFood and Drug Administration, HHS - Food & Beverage FDA 21 C.F.R. 175.300Dragoslav DzolicÎncă nu există evaluări
- Carboxylic Acids: Some Important Acids and Their Structural Formula'sDocument11 paginiCarboxylic Acids: Some Important Acids and Their Structural Formula'sKarthik SharmaÎncă nu există evaluări
- PTA Technology - FundamentalsDocument50 paginiPTA Technology - FundamentalsSarat Na NiÎncă nu există evaluări
- Daftar Obat Slow Moving Dan Ed Rawat Inap Maret 2021Document8 paginiDaftar Obat Slow Moving Dan Ed Rawat Inap Maret 2021Vima LadipaÎncă nu există evaluări
- Inserto Biorad Controles PDFDocument2 paginiInserto Biorad Controles PDFlenin_villalta67% (3)
- Polymers: Lecture 1 Unit-3a: Prepared by DR Arunima Nayak and DR Brij BhushanDocument2 paginiPolymers: Lecture 1 Unit-3a: Prepared by DR Arunima Nayak and DR Brij BhushanUtkarsh Singh0% (1)
- Puskesmas Citta: Pemerintah Kabupaten SoppengDocument12 paginiPuskesmas Citta: Pemerintah Kabupaten SoppengIsmach MizzleÎncă nu există evaluări
- MBM Home Quiz BEFORE MIDTERMDocument38 paginiMBM Home Quiz BEFORE MIDTERMbhagavan prasadÎncă nu există evaluări
- MODULE 7 Lesson1 - General Biology IDocument32 paginiMODULE 7 Lesson1 - General Biology IAlvin PaboresÎncă nu există evaluări