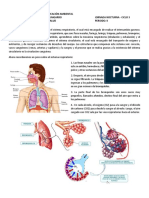
S-ar putea să vă placă și
- Fertirrigacion Cult Sin SueloDocument246 paginiFertirrigacion Cult Sin SueloSebastian Toronjo VazÎncă nu există evaluări
- Lixiviado de Lombris en Chile Habanero PDFDocument24 paginiLixiviado de Lombris en Chile Habanero PDFOmar Yavhe PsÎncă nu există evaluări
- Enfermedades y Plagas de La Papaya-Guia IlustradaDocument44 paginiEnfermedades y Plagas de La Papaya-Guia IlustradaEstrella LuceroÎncă nu există evaluări
- Manejo integrado de la polilla guatemalteca de la papa (Tecia solanivora, Povolny)De la EverandManejo integrado de la polilla guatemalteca de la papa (Tecia solanivora, Povolny)Încă nu există evaluări
- Cultivo de CempasúchilDocument7 paginiCultivo de CempasúchilMajo TorresÎncă nu există evaluări
- Manejo Integrado de PiñaDocument17 paginiManejo Integrado de PiñaKarina VargasÎncă nu există evaluări
- El Cultivo Del LitchiDocument7 paginiEl Cultivo Del LitchiRicardo Del Angel PoncianoÎncă nu există evaluări
- El Cultivo de CastañaDocument10 paginiEl Cultivo de CastañaRosali Ore Caballero0% (1)
- Enfermedades y Plagas de La Papaya en CordobaDocument5 paginiEnfermedades y Plagas de La Papaya en CordobaMANUEL MARTINEZÎncă nu există evaluări
- Cultivo de SesamoDocument18 paginiCultivo de SesamoGuilleGamer VA100% (1)
- CitoquiminaDocument6 paginiCitoquiminamarcelino canchari caceresÎncă nu există evaluări
- LitchiDocument2 paginiLitchiAna Virginia Sanchez Munguia0% (1)
- El Nopal Tunero en San Luis PotosíDocument39 paginiEl Nopal Tunero en San Luis PotosíMario RamosÎncă nu există evaluări
- El Cultivo Del Aguacate Razas y VariedadesDocument6 paginiEl Cultivo Del Aguacate Razas y VariedadesgusagroÎncă nu există evaluări
- Libro Simposio de Berries 2021Document112 paginiLibro Simposio de Berries 2021CinthiaLisbethGarciaZapataÎncă nu există evaluări
- Cultivo de MacaDocument21 paginiCultivo de Macajoshuadc89Încă nu există evaluări
- MILDIUDocument38 paginiMILDIUDiego Omar Blas CalizayaÎncă nu există evaluări
- Cultivo de BrocoliDocument21 paginiCultivo de BrocoliChristian Gilberto Cortez ChupinÎncă nu există evaluări
- Curso Naranja PDFDocument128 paginiCurso Naranja PDFeduadoÎncă nu există evaluări
- Trabajo Final de IsidroDocument46 paginiTrabajo Final de IsidroEdwin J. MartinezÎncă nu există evaluări
- Conferencia Ing. Eduardo Mejía VélezDocument28 paginiConferencia Ing. Eduardo Mejía VélezAndres F. VasquezÎncă nu există evaluări
- Manejo Agronomico en El Cultivo de La Quinua-IcaDocument18 paginiManejo Agronomico en El Cultivo de La Quinua-IcaJustin WatsonÎncă nu există evaluări
- Cana de Azucar - Maquinaria AgricolaDocument6 paginiCana de Azucar - Maquinaria AgricolaManuelRivasChilquilloÎncă nu există evaluări
- Tratamiento de Semillas PDFDocument79 paginiTratamiento de Semillas PDFLara Murrieta PraxedesÎncă nu există evaluări
- CIRUELADocument16 paginiCIRUELADavid Ernesto Mayen MilianÎncă nu există evaluări
- Importancia de La OlericulturaDocument13 paginiImportancia de La OlericulturaMaria Novelo CauichÎncă nu există evaluări
- Manual Fontagro Preparacion Pefiles de ProyectosDocument99 paginiManual Fontagro Preparacion Pefiles de ProyectosJuan José Aparicio Porres100% (1)
- APIOODocument35 paginiAPIOOCrhistiam Rudy Asqui SarazaÎncă nu există evaluări
- Antracnosis en MangoDocument24 paginiAntracnosis en Mangodaker12345Încă nu există evaluări
- ESCARIFICACIÓNDocument7 paginiESCARIFICACIÓNCarmelita Hcs RmrezÎncă nu există evaluări
- Evaluacion de Difente Densidad de Siembra en El Cultivo de Frejol en San Miguel de UrcuquíDocument18 paginiEvaluacion de Difente Densidad de Siembra en El Cultivo de Frejol en San Miguel de UrcuquíErika GualotoÎncă nu există evaluări
- Fenologia de La Frambuesa Roja Autumn Bliss en GDocument7 paginiFenologia de La Frambuesa Roja Autumn Bliss en GFernanda LopezÎncă nu există evaluări
- Taxonomia AguacateDocument3 paginiTaxonomia AguacateGeorginaÎncă nu există evaluări
- LechugaDocument2 paginiLechugaalexandraÎncă nu există evaluări
- 2.4 Produccion de Flores 4-Fusionado PDFDocument213 pagini2.4 Produccion de Flores 4-Fusionado PDFUFSFSDAJÎncă nu există evaluări
- Diagnostico Visual de Las PlantasDocument3 paginiDiagnostico Visual de Las PlantasGremberg Lopez100% (1)
- JicamaDocument12 paginiJicamaJaquelin NitoÎncă nu există evaluări
- La Siembra DirectaDocument10 paginiLa Siembra DirectaLeiner MelendezÎncă nu există evaluări
- El Cultivo de SojaDocument34 paginiEl Cultivo de SojaChi NoÎncă nu există evaluări
- Cultivo de ArvejaDocument26 paginiCultivo de ArvejaGrezia Coaquira RiosÎncă nu există evaluări
- Arrieta Quiñones Juan Carlos Carta Tecnologica Plantula de PimientoDocument3 paginiArrieta Quiñones Juan Carlos Carta Tecnologica Plantula de PimientoJudy H. AngelesÎncă nu există evaluări
- Condiciones Agroclimáticas - PaltoDocument2 paginiCondiciones Agroclimáticas - PaltoharrisonaqpÎncă nu există evaluări
- 5.1.1 Requerimientos Hidricos Del MangoDocument32 pagini5.1.1 Requerimientos Hidricos Del MangoOscar CotaÎncă nu există evaluări
- Bacteria 1Document16 paginiBacteria 1ASYSÎncă nu există evaluări
- Clavel TrabajoDocument31 paginiClavel TrabajoJean Franco ValdiviaÎncă nu există evaluări
- MIP Del RambutanDocument6 paginiMIP Del Rambutansandra100% (1)
- Expo Zanahoria y BetarragaDocument26 paginiExpo Zanahoria y BetarragaAbimael Sinche RamosÎncă nu există evaluări
- Virus Del Enrrollamiento de Las Hojas de La Papa (PLRV)Document7 paginiVirus Del Enrrollamiento de Las Hojas de La Papa (PLRV)JOHNBRAYHAN CANDIA CUTIPAÎncă nu există evaluări
- Cadena de Avena FINALDocument59 paginiCadena de Avena FINALmarx_hp04Încă nu există evaluări
- Manejo Agronómico Del RepolloDocument7 paginiManejo Agronómico Del RepolloJesús Alberto Sierra MorenoÎncă nu există evaluări
- 2488 - Folleto Super SorgoDocument56 pagini2488 - Folleto Super SorgoAlberto MaldonadoÎncă nu există evaluări
- Comercializacion Frijol CaupiDocument19 paginiComercializacion Frijol CaupiJosse Guerrero Vilela50% (2)
- Papaya 2019Document62 paginiPapaya 2019Nazly Yuliana Azabache MedinaÎncă nu există evaluări
- Tec Guia Manejo de MalezasDocument12 paginiTec Guia Manejo de MalezasCarolina MouraÎncă nu există evaluări
- Folleto ZarzamoraDocument35 paginiFolleto ZarzamoraAnonymous zmJVyN1SKVÎncă nu există evaluări
- Tesis-233 Ingeniería Agronómica - CD 635Document86 paginiTesis-233 Ingeniería Agronómica - CD 635Darcy GonzalezÎncă nu există evaluări
- FrijolDocument39 paginiFrijolUlises Gerardo Garcia ArmasÎncă nu există evaluări
- Enfermedades Solanaceas PDFDocument100 paginiEnfermedades Solanaceas PDFgatinha3350% (2)
- Infografía - Jose David LópezDocument2 paginiInfografía - Jose David LópezDaniela Garcés QuirogaÎncă nu există evaluări
- Infografía QuímicaDocument2 paginiInfografía QuímicaDaniela Garcés QuirogaÎncă nu există evaluări
- Bitácora Daniela Garcés 2191682Document1 paginăBitácora Daniela Garcés 2191682Daniela Garcés QuirogaÎncă nu există evaluări
- MORFOLOGIA Y FUNCION II Sistema NerviosoDocument56 paginiMORFOLOGIA Y FUNCION II Sistema NerviosoDaniela Garcés QuirogaÎncă nu există evaluări
- Protocolo de Prevención y Seguridad para El Manejo de Alimentos y Plazas de MercadoDocument2 paginiProtocolo de Prevención y Seguridad para El Manejo de Alimentos y Plazas de MercadoDaniela Garcés QuirogaÎncă nu există evaluări
- ELECTROLITOSDocument3 paginiELECTROLITOSDaniela Garcés QuirogaÎncă nu există evaluări
- Adaptaciones Fisiológicas Necesarias para El BuceoDocument33 paginiAdaptaciones Fisiológicas Necesarias para El BuceoMaria Luisa PuertaÎncă nu există evaluări
- Cap. 20 - Gasto Cardíaco, Retorno Venoso y Su Regulación PDFDocument8 paginiCap. 20 - Gasto Cardíaco, Retorno Venoso y Su Regulación PDFmaira ikesÎncă nu există evaluări
- CorazónDocument63 paginiCorazónVanessa Corona SalvadorÎncă nu există evaluări
- Guia Ciclo 3 Efectos Del Tabaco en La SaludDocument4 paginiGuia Ciclo 3 Efectos Del Tabaco en La SaludMaria Jose Villarreal MenaÎncă nu există evaluări
- Tipos de ShockDocument3 paginiTipos de ShockBrian96cortezÎncă nu există evaluări
- Anatomia Del CorazonDocument23 paginiAnatomia Del CorazonJesusAlejandroBernabeÎncă nu există evaluări
- Órganos LinfoidesDocument1 paginăÓrganos LinfoidesMareli M Damas Cicler100% (1)
- Exposic. Ciclo CardiacoDocument19 paginiExposic. Ciclo CardiacoHenrryPisfilÎncă nu există evaluări
- Nutrir El InteriorDocument6 paginiNutrir El InteriorJoan PozoÎncă nu există evaluări
- Cuaderno Cono 6 PrimariaDocument88 paginiCuaderno Cono 6 PrimariaRaquel ContrerasÎncă nu există evaluări
- Bloqueo Auriculoventricular Mobitz 2Document7 paginiBloqueo Auriculoventricular Mobitz 2Zharick vanessa Carrillo Dias100% (1)
- VerliannyDocument7 paginiVerliannygiovanny escalonaÎncă nu există evaluări
- Proyecto Cuerpo Humano y Los SistemasDocument16 paginiProyecto Cuerpo Humano y Los SistemasCarolina Sanchez Alonso67% (9)
- Coronari AsDocument31 paginiCoronari AsNely Andres MolinaÎncă nu există evaluări
- Carlos y Andres Sistema CirculatorioDocument20 paginiCarlos y Andres Sistema CirculatorioJUAN PABLO ACOSTA CUBILLOSÎncă nu există evaluări
- MATERNO INFANTIL Actualización 2022Document653 paginiMATERNO INFANTIL Actualización 2022Cynthia GutierrezÎncă nu există evaluări
- Fisiopatología Del ShockDocument30 paginiFisiopatología Del ShockYsabelÎncă nu există evaluări
- Test Hipertension Arterial ProfesoresDocument3 paginiTest Hipertension Arterial Profesoresfruben19Încă nu există evaluări
- Mudras Mag 8Document8 paginiMudras Mag 8Moises Cabello FernandezÎncă nu există evaluări
- ReflexologiaDocument23 paginiReflexologiapaola19Încă nu există evaluări
- Quiz - Escenario 3 - PRIMER BLOQUE-TEORICO - BIOLOGIA HUMANA - (GRUPO B01)Document8 paginiQuiz - Escenario 3 - PRIMER BLOQUE-TEORICO - BIOLOGIA HUMANA - (GRUPO B01)Lina Poveda0% (1)
- Derivados MesodermicosDocument9 paginiDerivados MesodermicosAnonymous C0v5OBY0TnÎncă nu există evaluări
- Atresia Pulmonar Con Tabique Interventricular IntegroDocument3 paginiAtresia Pulmonar Con Tabique Interventricular IntegroLuis Beltran MendozaÎncă nu există evaluări
- Listado Nominal de Gestantes 2018 Enero IiDocument4.379 paginiListado Nominal de Gestantes 2018 Enero IiVeronica Avila CozÎncă nu există evaluări
- Ficha Circulacion en El Ser HumanoDocument1 paginăFicha Circulacion en El Ser HumanoPaola LanzianoÎncă nu există evaluări
- Ritmo SinusalDocument3 paginiRitmo SinusalPaulina Rosas GleasonÎncă nu există evaluări
- Actividades Fisicas y Deportivas Bloque I, Ii, Iii Semestre A, Ana LiliaDocument116 paginiActividades Fisicas y Deportivas Bloque I, Ii, Iii Semestre A, Ana LiliaIMELDA HERNANDEZ GARCIA100% (1)
- Subtema 5 Nutric. AnimalesDocument2 paginiSubtema 5 Nutric. AnimalesDiana Caroline Arevalo VelaÎncă nu există evaluări
- Corazon GeneralidadesDocument52 paginiCorazon GeneralidadesTannyta CordovaÎncă nu există evaluări
- Anatomía Del Corazón: Tiene La Forma de Un Cono o Una Pirámide. en Él Pueden ReconocerseDocument13 paginiAnatomía Del Corazón: Tiene La Forma de Un Cono o Una Pirámide. en Él Pueden ReconocerseMarÎncă nu există evaluări