S-ar putea să vă placă și
- The Role of MuscleDocument5 paginiThe Role of MuscleMuhammad FaiqÎncă nu există evaluări
- Exogenous Proteases For Meat TenderizationDocument89 paginiExogenous Proteases For Meat TenderizationMuhammad FaiqÎncă nu există evaluări
- Exogenous Proteases For Meat TenderizationDocument89 paginiExogenous Proteases For Meat TenderizationMuhammad FaiqÎncă nu există evaluări
- DPPH AssayDocument1 paginăDPPH AssayMuhammad FaiqÎncă nu există evaluări
- Bound Phenolics in Foods, A ReviewDocument10 paginiBound Phenolics in Foods, A ReviewMuhammad FaiqÎncă nu există evaluări
- Microsoft Word - Making SolutionsDocument5 paginiMicrosoft Word - Making SolutionsMuhammad FaiqÎncă nu există evaluări
- Dog Rose and Pomegranate Extracts As Agents To Control Enzymatic BrowningDocument7 paginiDog Rose and Pomegranate Extracts As Agents To Control Enzymatic BrowningMuhammad FaiqÎncă nu există evaluări
- A Buffer Solution For ColorimetricDocument5 paginiA Buffer Solution For ColorimetricMuhammad FaiqÎncă nu există evaluări
- Antioxidant Activity and Phenolic Compounds in 32 Selected Herbs PDFDocument10 paginiAntioxidant Activity and Phenolic Compounds in 32 Selected Herbs PDFMuhammad FaiqÎncă nu există evaluări
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Elpodereso Case AnalysisDocument3 paginiElpodereso Case AnalysisUsama17100% (2)
- Statistics and Probability: Quarter 4 - (Week 6)Document8 paginiStatistics and Probability: Quarter 4 - (Week 6)Jessa May MarcosÎncă nu există evaluări
- BJAS - Volume 5 - Issue Issue 1 Part (2) - Pages 275-281Document7 paginiBJAS - Volume 5 - Issue Issue 1 Part (2) - Pages 275-281Vengky UtamiÎncă nu există evaluări
- WHO Guidelines For Drinking Water: Parameters Standard Limits As Per WHO Guidelines (MG/L)Document3 paginiWHO Guidelines For Drinking Water: Parameters Standard Limits As Per WHO Guidelines (MG/L)114912Încă nu există evaluări
- FpsecrashlogDocument19 paginiFpsecrashlogtim lokÎncă nu există evaluări
- Building and Structural Construction N6 T1 2024 T2Document9 paginiBuilding and Structural Construction N6 T1 2024 T2FranceÎncă nu există evaluări
- Pipe Cleaner Lesson PlanDocument2 paginiPipe Cleaner Lesson PlanTaylor FranklinÎncă nu există evaluări
- Biological Assets Sample ProblemsDocument4 paginiBiological Assets Sample ProblemsKathleenÎncă nu există evaluări
- CS8CHP EletricalDocument52 paginiCS8CHP EletricalCristian ricardo russoÎncă nu există evaluări
- Wwii TictactoeDocument2 paginiWwii Tictactoeapi-557780348Încă nu există evaluări
- Low Budget Music Promotion and PublicityDocument41 paginiLow Budget Music Promotion and PublicityFola Folayan100% (3)
- Hydro Electric Fire HistoryDocument3 paginiHydro Electric Fire HistorygdmurfÎncă nu există evaluări
- ETAP Power Station ErrorDocument5 paginiETAP Power Station ErroryogacruiseÎncă nu există evaluări
- Electronic Spin Inversion: A Danger To Your HealthDocument4 paginiElectronic Spin Inversion: A Danger To Your Healthambertje12Încă nu există evaluări
- Draft PDFDocument166 paginiDraft PDFashwaq000111Încă nu există evaluări
- VMP 930 Veterinary Parasitology: Paragonimus KellicottiDocument63 paginiVMP 930 Veterinary Parasitology: Paragonimus KellicottiRenien Khim BahayaÎncă nu există evaluări
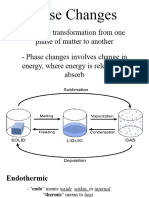
- General Chemistry 2 Q1 Lesson 5 Endothermic and Exotheric Reaction and Heating and Cooling CurveDocument19 paginiGeneral Chemistry 2 Q1 Lesson 5 Endothermic and Exotheric Reaction and Heating and Cooling CurveJolo Allexice R. PinedaÎncă nu există evaluări
- Practising Modern English For Life Sciences Students Caiet Exercitii-1Document77 paginiPractising Modern English For Life Sciences Students Caiet Exercitii-1Robert BobiaÎncă nu există evaluări
- Open Letter To Hon. Nitin Gadkari On Pothole Problem On National and Other Highways in IndiaDocument3 paginiOpen Letter To Hon. Nitin Gadkari On Pothole Problem On National and Other Highways in IndiaProf. Prithvi Singh KandhalÎncă nu există evaluări
- Alkosign Product CatalogeDocument20 paginiAlkosign Product CatalogeShree AgrawalÎncă nu există evaluări
- Financial Management 2E: Rajiv Srivastava - Dr. Anil Misra Solutions To Numerical ProblemsDocument5 paginiFinancial Management 2E: Rajiv Srivastava - Dr. Anil Misra Solutions To Numerical ProblemsParesh ShahÎncă nu există evaluări
- Ducted Split ACsDocument31 paginiDucted Split ACsHammadZaman100% (1)
- Recruitment SelectionDocument11 paginiRecruitment SelectionMOHAMMED KHAYYUMÎncă nu există evaluări
- Grid Pattern PortraitDocument8 paginiGrid Pattern PortraitEmma FravigarÎncă nu există evaluări
- Question Answers of Chapter 13 Class 5Document6 paginiQuestion Answers of Chapter 13 Class 5SuvashreePradhanÎncă nu există evaluări
- MME 52106 - Optimization in Matlab - NN ToolboxDocument14 paginiMME 52106 - Optimization in Matlab - NN ToolboxAdarshÎncă nu există evaluări
- Espree I Class Korr3Document22 paginiEspree I Class Korr3hgaucherÎncă nu există evaluări
- Lesson Plan For Implementing NETSDocument5 paginiLesson Plan For Implementing NETSLisa PizzutoÎncă nu există evaluări
- 2-1. Drifting & Tunneling Drilling Tools PDFDocument9 pagini2-1. Drifting & Tunneling Drilling Tools PDFSubhash KediaÎncă nu există evaluări
- Virtual Assets Act, 2022Document18 paginiVirtual Assets Act, 2022Rapulu UdohÎncă nu există evaluări