S-ar putea să vă placă și
- Le Sexe Pour Les NulsDocument1.355 paginiLe Sexe Pour Les Nulsjimmy100% (1)
- Développement végétal: Les Grands Articles d'UniversalisDe la EverandDéveloppement végétal: Les Grands Articles d'UniversalisÎncă nu există evaluări
- Appareil génital: Les Grands Articles d'UniversalisDe la EverandAppareil génital: Les Grands Articles d'UniversalisÎncă nu există evaluări
- 778 Evolution Des Classifications BotaniquesDocument50 pagini778 Evolution Des Classifications BotaniquesHacerMellalÎncă nu există evaluări
- Polymorphisme en biologie: Les Grands Articles d'UniversalisDe la EverandPolymorphisme en biologie: Les Grands Articles d'UniversalisÎncă nu există evaluări
- TP N 2 Etude de La Cellule Animale Et VegetaleDocument4 paginiTP N 2 Etude de La Cellule Animale Et Vegetalemaysoun100% (2)
- TP N 1 La MicroscopieDocument5 paginiTP N 1 La MicroscopieJoujou Hadjine100% (2)
- La SpermatogeneseDocument21 paginiLa SpermatogeneseFaculté De Médecine Béchar100% (2)
- Biologie Végétale Exercices Corrigé 01Document3 paginiBiologie Végétale Exercices Corrigé 01maysoun100% (1)
- La Reproduction de L'homme Et L'éducation À La Sexualité CM2Document18 paginiLa Reproduction de L'homme Et L'éducation À La Sexualité CM2lezardkingÎncă nu există evaluări
- (Aristote) de La G?n?ration Des AnimauxDocument461 pagini(Aristote) de La G?n?ration Des AnimauxFabio Prado de FreitasÎncă nu există evaluări
- Cours Origine Du Génotype Des IndividusDocument5 paginiCours Origine Du Génotype Des IndividusjoanaÎncă nu există evaluări
- TD Gen Pop Agro2Document6 paginiTD Gen Pop Agro2Freenel BossaÎncă nu există evaluări
- Selection Et Amelioration Genetique Chap 1Document6 paginiSelection Et Amelioration Genetique Chap 1Saida AdjemiÎncă nu există evaluări
- I3 Stabilité Et Variabilité Du Génome Et Évolution: 1ère PARTIEDocument17 paginiI3 Stabilité Et Variabilité Du Génome Et Évolution: 1ère PARTIEloveliineÎncă nu există evaluări
- Stabilité Et Variabilité Des Génômes Et ÉvolutionDocument6 paginiStabilité Et Variabilité Des Génômes Et ÉvolutionHellie WrightÎncă nu există evaluări
- Le Brassage Génétique Par La Méiose Et La FécondationDocument15 paginiLe Brassage Génétique Par La Méiose Et La Fécondationbouchakour meryemÎncă nu există evaluări
- Doc 03Document20 paginiDoc 03hanane boutajÎncă nu există evaluări
- Diplo - Liaison Genetique Et Cartographie GenetiqueDocument11 paginiDiplo - Liaison Genetique Et Cartographie GenetiqueAdmi BraccÎncă nu există evaluări
- Cas Particuliers Du MendélismeDocument3 paginiCas Particuliers Du Mendélismerolling stoneÎncă nu există evaluări
- 8 TP7 Grps SG CorrecDocument4 pagini8 TP7 Grps SG CorrecibrahimÎncă nu există evaluări
- Chap - 3 SVT TDDocument5 paginiChap - 3 SVT TDRebecca MoulackaÎncă nu există evaluări
- HAV504B - Chapitre 1Document7 paginiHAV504B - Chapitre 1Ylona RamondÎncă nu există evaluări
- Option de Génétique BIO 60Document25 paginiOption de Génétique BIO 60lieu duÎncă nu există evaluări
- Ségrégation D'un Couple D'allèlesDocument55 paginiSégrégation D'un Couple D'allèlesYves-Nathan Tian BiÎncă nu există evaluări
- Haplophase - Différence Due À Un Seul Gène 1Document5 paginiHaplophase - Différence Due À Un Seul Gène 1Admi BraccÎncă nu există evaluări
- Ilovepdf Merged (2) RemovedDocument12 paginiIlovepdf Merged (2) RemovedMa NouÎncă nu există evaluări
- ExtraitDocument10 paginiExtraitSoumanaÎncă nu există evaluări
- Génétique SynthèseDocument55 paginiGénétique SynthèseArnaud LiebinÎncă nu există evaluări
- $R7FKSLTDocument10 pagini$R7FKSLToumaimabenyoussof03Încă nu există evaluări
- POLY Genetique DCEM1 1Document54 paginiPOLY Genetique DCEM1 1mouha4100% (1)
- Chap 1.1 - Origine Du Génotype Des IndividusDocument4 paginiChap 1.1 - Origine Du Génotype Des IndividusicemerproÎncă nu există evaluări
- Notions Amélioration PlantesDocument4 paginiNotions Amélioration PlantesPatrick EbaÎncă nu există evaluări
- Devoir Bio Février 2014 TSEXPDocument3 paginiDevoir Bio Février 2014 TSEXPSoumana SogobaÎncă nu există evaluări
- Seq 10 Cours Génétique RedigéDocument6 paginiSeq 10 Cours Génétique RedigéLadetÎncă nu există evaluări
- Cours 2Document82 paginiCours 2imenbmc1Încă nu există evaluări
- Items de Biologie (2013 À 2018)Document15 paginiItems de Biologie (2013 À 2018)pathenrscodeÎncă nu există evaluări
- UAA 8 - chp2 - Prolongements - Mendel - 2020 - Partie2Document8 paginiUAA 8 - chp2 - Prolongements - Mendel - 2020 - Partie2céline prielsÎncă nu există evaluări
- Cours de Tle D - Séquence 5Document8 paginiCours de Tle D - Séquence 5Tite DembeleÎncă nu există evaluări
- Pharm2an16 Genetique-DihybridismeDocument60 paginiPharm2an16 Genetique-Dihybridismeyassine rekikÎncă nu există evaluări
- Innovations Génétiques - WikipédiaDocument23 paginiInnovations Génétiques - WikipédiaKaton dit abdramane CoulibalyÎncă nu există evaluări
- Document PDF 4Document1 paginăDocument PDF 4pzÎncă nu există evaluări
- Localisation Des GènesDocument27 paginiLocalisation Des Gènesjulien KoudouÎncă nu există evaluări
- Monohybridisme Chez Les DiploïdesDocument8 paginiMonohybridisme Chez Les DiploïdesImene NamousÎncă nu există evaluări
- 18 TS 1A1 LWPDocument12 pagini18 TS 1A1 LWPfardadi rachidÎncă nu există evaluări
- 01 - Le Brassage Génétique Et Sa Contribution À La Diversité GénétiqueDocument7 pagini01 - Le Brassage Génétique Et Sa Contribution À La Diversité GénétiqueClohtaire Le NiduÎncă nu există evaluări
- Pour Réussir Les Exercices deDocument4 paginiPour Réussir Les Exercices deOumou Khaïry Diop100% (3)
- Chapitre 2Document49 paginiChapitre 2Rahma HosniÎncă nu există evaluări
- Chapitre L'origine de La Diversité Génétique Des IndividusDocument20 paginiChapitre L'origine de La Diversité Génétique Des IndividusSriÎncă nu există evaluări
- Localisation Des GenesDocument27 paginiLocalisation Des GenesDenisco NkemgneÎncă nu există evaluări
- Cartes GénétiquesDocument53 paginiCartes GénétiquesFaiza HÎncă nu există evaluări
- Chap 1 SVT Termnale SDocument6 paginiChap 1 SVT Termnale SghamguiÎncă nu există evaluări
- Derive Genetique Selection SpeciationDocument3 paginiDerive Genetique Selection SpeciationmaenapouqetÎncă nu există evaluări
- Brassage GénétiqueDocument5 paginiBrassage Génétiquele sage100% (1)
- Cours Génétique Des HaploidesDocument33 paginiCours Génétique Des Haploidesbenamernessrine8Încă nu există evaluări
- Exos Genetique Des PopDocument92 paginiExos Genetique Des PopFarrouha Haddad100% (1)
- Cours Préparatoire Biologie Cours 9 Dia MoodleDocument86 paginiCours Préparatoire Biologie Cours 9 Dia MoodleRym ZakraouiÎncă nu există evaluări
- MeaamriDocument16 paginiMeaamriMaroua DellidjÎncă nu există evaluări
- Activite 3 Les Mecanismes Evolutifs Partie B La Derive GenetiqueDocument2 paginiActivite 3 Les Mecanismes Evolutifs Partie B La Derive Genetiquefaouziaya605Încă nu există evaluări
- Evolution MécanismesDocument4 paginiEvolution Mécanismessassitakwa07Încă nu există evaluări
- S SVT Obligatoire 2019 Polynesie CorrigeDocument8 paginiS SVT Obligatoire 2019 Polynesie CorrigeKahina MallilÎncă nu există evaluări
- Chapitre 1 1 L Origine Du Ge Notype Des Individus Cours e Le VeDocument6 paginiChapitre 1 1 L Origine Du Ge Notype Des Individus Cours e Le VeIsabel RamírezÎncă nu există evaluări
- TSPE - T1 - C1 - Notions 3Document2 paginiTSPE - T1 - C1 - Notions 3laure.c.lucetteÎncă nu există evaluări
- Ed Genetique CorrigeDocument51 paginiEd Genetique CorrigetafarelantenovÎncă nu există evaluări
- Chapitres 3 & 4Document17 paginiChapitres 3 & 4Njiasse MaazouÎncă nu există evaluări
- Modes de Transmission Hereditaire Des Caracteres MonofactorielsDocument9 paginiModes de Transmission Hereditaire Des Caracteres MonofactorielsMarietteÎncă nu există evaluări
- 656734fb55592 - Énoncé - Série 12 - DihybridismeDocument4 pagini656734fb55592 - Énoncé - Série 12 - Dihybridismesekrafinidhal0% (1)
- CH4 - Mécanismes Évolutifs Et BiodiversitéDocument3 paginiCH4 - Mécanismes Évolutifs Et BiodiversitéjoseethermidorÎncă nu există evaluări
- Pré Analytique GEHT 2018Document2 paginiPré Analytique GEHT 2018maysounÎncă nu există evaluări
- TP 011Document2 paginiTP 011maysounÎncă nu există evaluări
- TP 0 Présentation Du LaboratoireDocument7 paginiTP 0 Présentation Du LaboratoiremaysounÎncă nu există evaluări
- Pré Analytique GEHT 2018Document2 paginiPré Analytique GEHT 2018maysounÎncă nu există evaluări
- TP 0 Présentation Du LaboratoireDocument7 paginiTP 0 Présentation Du LaboratoiremaysounÎncă nu există evaluări
- Extrait - Methodes de Creation de Varietes en AmeliorDocument20 paginiExtrait - Methodes de Creation de Varietes en Ameliormirou lbÎncă nu există evaluări
- Des SubstancesDocument1 paginăDes SubstancesmaysounÎncă nu există evaluări
- Biologie Végétale Résumé 02Document7 paginiBiologie Végétale Résumé 02maysounÎncă nu există evaluări
- Biologie Végétale Résumé 04 PDFDocument5 paginiBiologie Végétale Résumé 04 PDFmaysounÎncă nu există evaluări
- IntroductionDocument1 paginăIntroductionmaysounÎncă nu există evaluări
- Biologie Végétale Résumé 01Document4 paginiBiologie Végétale Résumé 01maysounÎncă nu există evaluări
- Biologie Végétale Résumé 03Document4 paginiBiologie Végétale Résumé 03maysounÎncă nu există evaluări
- ClassificationDocument10 paginiClassificationmaysounÎncă nu există evaluări
- Biologie Vegetale 2015Document1 paginăBiologie Vegetale 2015maysounÎncă nu există evaluări
- ArchitectureDocument1 paginăArchitecturemaysounÎncă nu există evaluări
- Corrigé Type EMD de Biologie AnimaleDocument2 paginiCorrigé Type EMD de Biologie Animalemaysoun100% (1)
- TD N 2 Cytologie Synthese ProteiqueDocument3 paginiTD N 2 Cytologie Synthese ProteiquemaysounÎncă nu există evaluări
- TD N 2 Cytologie Synthese ProteiqueDocument3 paginiTD N 2 Cytologie Synthese ProteiquemaysounÎncă nu există evaluări
- TD N1 Le Cycle CellulaireDocument6 paginiTD N1 Le Cycle Cellulairemaysoun100% (1)
- TéléchargementDocument5 paginiTéléchargementmaysounÎncă nu există evaluări
- GenèseDocument1 paginăGenèsemaysounÎncă nu există evaluări
- Corrige Rattrapage-Biologie-animale-2015Document2 paginiCorrige Rattrapage-Biologie-animale-2015maysounÎncă nu există evaluări
- TP N 3 Echanges CellulairesDocument4 paginiTP N 3 Echanges Cellulairesmaysoun100% (1)
- TP N 4 ParamecieDocument4 paginiTP N 4 ParameciemaysounÎncă nu există evaluări
- TP N 3 Echanges CellulairesDocument4 paginiTP N 3 Echanges Cellulairesmaysoun100% (1)
- TP N 4 ParamecieDocument4 paginiTP N 4 ParameciemaysounÎncă nu există evaluări
- كتاب العلوم للسنة السابعة 7D PDFDocument209 paginiكتاب العلوم للسنة السابعة 7D PDFLeinad BarrackÎncă nu există evaluări
- 2nd - Chp3 - TP1 (Correction)Document4 pagini2nd - Chp3 - TP1 (Correction)ademsaai4Încă nu există evaluări
- 6sdevoir de Synthèse N°1Document6 pagini6sdevoir de Synthèse N°1Bilel Amri0% (1)
- 5 16 1 PBDocument5 pagini5 16 1 PBOum BanatÎncă nu există evaluări
- (Chapitre 2 Organisation Fonctionnelle Des Systèmes Reproducteurs) PDFDocument14 pagini(Chapitre 2 Organisation Fonctionnelle Des Systèmes Reproducteurs) PDFSamra LadjelÎncă nu există evaluări
- FSH FSH: Exploration Des Troubles PubertairesDocument3 paginiFSH FSH: Exploration Des Troubles PubertairesKhawla BelkharchoucheÎncă nu există evaluări
- Actualite de Reproduction DromadaireDocument8 paginiActualite de Reproduction DromadaireBOUTHEINA BOUHALASSAÎncă nu există evaluări
- De La Fleur A La GraineDocument5 paginiDe La Fleur A La Graineamar4430Încă nu există evaluări
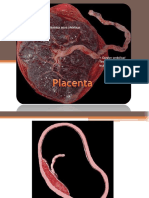
- PlacentaDocument27 paginiPlacentaحامدي يعقوبÎncă nu există evaluări
- BacDocument77 paginiBacSouhail El AsferÎncă nu există evaluări
- Pathologies Des Spécialités: Médicales Et Soins InfirmiersDocument15 paginiPathologies Des Spécialités: Médicales Et Soins InfirmiersouaskokmohamedÎncă nu există evaluări
- Corrige Type Harmonise Des Trois Centres de Correction Du Bac Ii 2022 - SVT Series A4-C-D Office Du BacDocument10 paginiCorrige Type Harmonise Des Trois Centres de Correction Du Bac Ii 2022 - SVT Series A4-C-D Office Du BacBilale RaouphÎncă nu există evaluări
- Page 32 of 551Document1 paginăPage 32 of 551Mohammed ASÎncă nu există evaluări
- La Stérilité FéminineDocument17 paginiLa Stérilité FéminineSofiane BousifeÎncă nu există evaluări
- Exercice SVTDocument5 paginiExercice SVTcoumouyeÎncă nu există evaluări
- ACTIVITE NOAM M 3ea LDHDocument3 paginiACTIVITE NOAM M 3ea LDHnm mrchwkÎncă nu există evaluări
- Abies Bornmuelleriana Mattf. Syn. Abies Equi-Trojani (Asch. & Sint. Ex Boiss.) Mattf. Sapin de Bornmüller, Sapin de La Mer NoireDocument6 paginiAbies Bornmuelleriana Mattf. Syn. Abies Equi-Trojani (Asch. & Sint. Ex Boiss.) Mattf. Sapin de Bornmüller, Sapin de La Mer NoireIsembockÎncă nu există evaluări
- CH 1 - TP 2Document4 paginiCH 1 - TP 2anissaattoumani1234Încă nu există evaluări
- Note de CoursDocument25 paginiNote de Courselena.rids135Încă nu există evaluări
- La Régulation de La Fonction Reproductrice - Réviser - Concours Paramédicaux - Sanitaire Et Social - Cap ConcoursDocument4 paginiLa Régulation de La Fonction Reproductrice - Réviser - Concours Paramédicaux - Sanitaire Et Social - Cap ConcoursKhaled JberiÎncă nu există evaluări
- TD Embryologie Organes Genitaux Masculins Et SpermatogeneseDocument3 paginiTD Embryologie Organes Genitaux Masculins Et Spermatogenesejulien KoudouÎncă nu există evaluări
- Benin 2022 Aperçu Des IndicateursDocument2 paginiBenin 2022 Aperçu Des IndicateursNaili Bourhane Aboudou RahimeÎncă nu există evaluări
- Presentation Du SiegeDocument8 paginiPresentation Du SiegeNătă ĈhăÎncă nu există evaluări
- 07 La-FleurDocument5 pagini07 La-FleurAriaÎncă nu există evaluări
- Cours ReproductionDocument7 paginiCours ReproductionBen MabÎncă nu există evaluări
- 371 - Concours Doctorat LMD 2012-2013-VFDocument14 pagini371 - Concours Doctorat LMD 2012-2013-VFzaki_b6Încă nu există evaluări