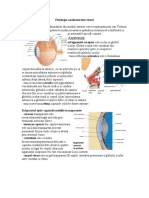
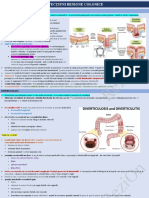
S-ar putea să vă placă și
- Sucul GastricDocument7 paginiSucul GastricAndreea MarinÎncă nu există evaluări
- Curs 5 - TrombocitulDocument24 paginiCurs 5 - TrombocitulNihilobstatÎncă nu există evaluări
- Curs 9 HistoDocument84 paginiCurs 9 HistoDan MunteanuÎncă nu există evaluări
- EritrociteleDocument13 paginiEritrociteleAdrian98Încă nu există evaluări
- HormoniDocument6 paginiHormoniLigia HumeniucÎncă nu există evaluări
- Curs 9 Metabolismul BilirubineiDocument32 paginiCurs 9 Metabolismul BilirubineiAndone Marlena100% (1)
- LP 11 - Hemostaza PrimaraDocument6 paginiLP 11 - Hemostaza PrimaraPetronela MateoiuÎncă nu există evaluări
- Fiziologia Analizatorului VizualDocument2 paginiFiziologia Analizatorului VizualPopa CarmenÎncă nu există evaluări
- Grile Microbiologie An 2 .. MGDocument13 paginiGrile Microbiologie An 2 .. MGAlex PomanaÎncă nu există evaluări
- Test General de BiologieDocument5 paginiTest General de BiologieAvillaÎncă nu există evaluări
- 2016 - Taus - Teste de Toxicologie ClinicaDocument229 pagini2016 - Taus - Teste de Toxicologie ClinicaAnonymous IOrWuyVQS1Încă nu există evaluări
- SangeleDocument11 paginiSangeleSofica PetronelaÎncă nu există evaluări
- LP I-4 21-43Document40 paginiLP I-4 21-43Ileana IoanaÎncă nu există evaluări
- Glandele-Suprarenale PatologieDocument32 paginiGlandele-Suprarenale PatologieVadimMorariÎncă nu există evaluări
- CURS 11 Metabolismul Lipidelor 1Document60 paginiCURS 11 Metabolismul Lipidelor 1waseemÎncă nu există evaluări
- Sistemul Nervos Vegetativ - COLINERGICDocument14 paginiSistemul Nervos Vegetativ - COLINERGICMihai MarinÎncă nu există evaluări
- Curs 11 Grupe SanguineDocument4 paginiCurs 11 Grupe SanguineElena Caranica100% (1)
- Hemoglobin ADocument32 paginiHemoglobin AAnda PopaÎncă nu există evaluări
- HematopoiezaDocument6 paginiHematopoiezaDaiana CiobanuÎncă nu există evaluări
- Subiecte ImunologieDocument116 paginiSubiecte ImunologieDiana IlieÎncă nu există evaluări
- LP 4 FiziopatologieDocument6 paginiLP 4 Fiziopatologiecmvio100% (1)
- Imunologie Text1Document469 paginiImunologie Text1Vra4100% (3)
- Numaratoarea Eritrocitelor, Trombocitelor Si Leucocitelor - LPDocument32 paginiNumaratoarea Eritrocitelor, Trombocitelor Si Leucocitelor - LPAna-MariaStanilaÎncă nu există evaluări
- Membrana CelularaDocument77 paginiMembrana CelularaCătălina RaicuÎncă nu există evaluări
- Tulbură-Rile Metabo - Lismului GlucidicDocument33 paginiTulbură-Rile Metabo - Lismului GlucidicCaletnic AlionaÎncă nu există evaluări
- Leucemia Granulocitara CronicaDocument31 paginiLeucemia Granulocitara Cronicanistor97Încă nu există evaluări
- ANEMIIDocument22 paginiANEMIIStefania NicolaeÎncă nu există evaluări
- Polipi - Cancer Colo-RectalDocument6 paginiPolipi - Cancer Colo-RectalVerinceanu AlexandruÎncă nu există evaluări
- Biosinteza AminoacizilorDocument2 paginiBiosinteza AminoacizilorIonut NealcosÎncă nu există evaluări
- Echinococcus GranulosusDocument18 paginiEchinococcus GranulosusXi ShiÎncă nu există evaluări
- 10 Metabolismul Hemoglobinei PrezentareDocument16 pagini10 Metabolismul Hemoglobinei PrezentareMariana Vesa100% (1)
- Referat HematologieDocument20 paginiReferat HematologieEugenia CosovanÎncă nu există evaluări
- Vitamina KDocument15 paginiVitamina KAlexandra MateiÎncă nu există evaluări
- Hiv SidaDocument3 paginiHiv Sidadoc buc100% (1)
- Clostridiile LPDocument8 paginiClostridiile LPCornelia HristiaÎncă nu există evaluări
- Subiecte Examen Fiziologie MG I SEM II SibiuDocument4 paginiSubiecte Examen Fiziologie MG I SEM II Sibiumaria_fratila_2Încă nu există evaluări
- PinocitozaDocument15 paginiPinocitozaIonut Bojor100% (1)
- FIZIOLOGIE Sangele2Document63 paginiFIZIOLOGIE Sangele2Mihaela VadanaÎncă nu există evaluări
- TesteDocument3 paginiTesteViolina CîmpanuÎncă nu există evaluări
- Test Grila Embriologie - OdtDocument4 paginiTest Grila Embriologie - OdtRotaru Lacramioara OanaÎncă nu există evaluări
- Fiziopatologie LaboratorDocument214 paginiFiziopatologie Laboratorp3tru_mzqÎncă nu există evaluări
- Hematologie - REZOLVATE PDFDocument5 paginiHematologie - REZOLVATE PDFRamona FlorinaÎncă nu există evaluări
- Anul 1 Semiologie Medicala - Curs 12Document9 paginiAnul 1 Semiologie Medicala - Curs 12Valérie CharisÎncă nu există evaluări
- 6 HematopoezaDocument8 pagini6 HematopoezaValentin MoisăÎncă nu există evaluări
- Hematopoieza Prenatala (Etape)Document1 paginăHematopoieza Prenatala (Etape)Ïulia LøÎncă nu există evaluări
- Excitabilitatea - BiofizicaDocument21 paginiExcitabilitatea - Biofizicamarius_inter881956Încă nu există evaluări
- Explorarea Metabolismului ProtidicDocument6 paginiExplorarea Metabolismului ProtidicRusu Stefan100% (1)
- Teste 2 Rom. (2008)Document21 paginiTeste 2 Rom. (2008)Rebeca SuruceanuÎncă nu există evaluări
- Coloratia VitalaDocument3 paginiColoratia Vitalalalu15Încă nu există evaluări
- Mecanismul de Actiune Al HormonilorDocument7 paginiMecanismul de Actiune Al HormonilorMaria Lungu100% (1)
- Curs 8 RenalDocument22 paginiCurs 8 RenalArwen MyÎncă nu există evaluări
- Grile HematologieDocument9 paginiGrile HematologieDana NițăÎncă nu există evaluări
- 3chimia, Patochimia ProteinelorDocument61 pagini3chimia, Patochimia ProteinelorSpinu LiliaÎncă nu există evaluări
- GlicemiaDocument13 paginiGlicemiaAnonymous b8pnWJM100% (1)
- Fiziologia Muschiului - 2019Document43 paginiFiziologia Muschiului - 2019Narcis MessiÎncă nu există evaluări
- 2018-2019 - Curs 5 FicatDocument56 pagini2018-2019 - Curs 5 FicatXerox medÎncă nu există evaluări
- Curs 12 Hemul Degradare Si BiosintezaDocument16 paginiCurs 12 Hemul Degradare Si BiosintezaMarkus ZahndÎncă nu există evaluări
- Tema: Producerea Și Metabolismul Bilirubinei: Efectuat: Șișianu Teodor, M1515Document14 paginiTema: Producerea Și Metabolismul Bilirubinei: Efectuat: Șișianu Teodor, M1515Teodor ŞişianuÎncă nu există evaluări
- Pigmentii BiliariDocument2 paginiPigmentii BiliarimiaubooksÎncă nu există evaluări
- Fiziologia FicatuluiDocument7 paginiFiziologia FicatuluiMaria FrancescaÎncă nu există evaluări
- Bilirubin ADocument4 paginiBilirubin AAlexandra LacatusuÎncă nu există evaluări
- Bilirubin ADocument5 paginiBilirubin AIrina PurlucaÎncă nu există evaluări
- Infecții Cu Transmitere SexualăDocument4 paginiInfecții Cu Transmitere SexualăVerinceanu AlexandruÎncă nu există evaluări
- ANATOMIEDocument1 paginăANATOMIEVerinceanu AlexandruÎncă nu există evaluări
- Intestin Gros - OcluzieDocument2 paginiIntestin Gros - OcluzieVerinceanu AlexandruÎncă nu există evaluări
- Managementul Reacțiilor Alergice in Unitatea de Primiri Urgente Articoloriginal 2016 RMUDocument6 paginiManagementul Reacțiilor Alergice in Unitatea de Primiri Urgente Articoloriginal 2016 RMUVerinceanu AlexandruÎncă nu există evaluări
- Volvulus de ColonDocument1 paginăVolvulus de ColonVerinceanu AlexandruÎncă nu există evaluări
- 11.8 Ocluziile Ale Ghid de StudiuDocument16 pagini11.8 Ocluziile Ale Ghid de StudiumateideniÎncă nu există evaluări
- ANATOMIEDocument3 paginiANATOMIEVerinceanu AlexandruÎncă nu există evaluări
- Colon - Pseudo-Ocluzie AcutăDocument2 paginiColon - Pseudo-Ocluzie AcutăVerinceanu AlexandruÎncă nu există evaluări
- Anexa I.1 PDFDocument1 paginăAnexa I.1 PDFRaluca-Andreea IgnatÎncă nu există evaluări
- Agentii de Voiaj CFR Actualizare Oficios 2018 V2 21 FebDocument3 paginiAgentii de Voiaj CFR Actualizare Oficios 2018 V2 21 FebClaudiu PopescuÎncă nu există evaluări
- Afecțiuni Benigne ColoniceDocument5 paginiAfecțiuni Benigne ColoniceVerinceanu AlexandruÎncă nu există evaluări
- Fisa Practica Anul MG 5Document2 paginiFisa Practica Anul MG 5Verinceanu AlexandruÎncă nu există evaluări
- 2019-20120 Orar Centralizator Clase Since 16.09.2019 LUNGADocument1 pagină2019-20120 Orar Centralizator Clase Since 16.09.2019 LUNGAVerinceanu AlexandruÎncă nu există evaluări
- Specialitati Deficitare2019Document1 paginăSpecialitati Deficitare2019Oana SmarandaÎncă nu există evaluări
- Coordonatori BRASOVDocument3 paginiCoordonatori BRASOVVerinceanu AlexandruÎncă nu există evaluări
- Formular 7 CERERE DE DESCHIDERE CONT ADITIONALDocument1 paginăFormular 7 CERERE DE DESCHIDERE CONT ADITIONALVerinceanu AlexandruÎncă nu există evaluări
- Rasp SPDocument20 paginiRasp SPVerinceanu AlexandruÎncă nu există evaluări
- Orar Recuperare 30.04.2018 Fiecare ProfesorDocument66 paginiOrar Recuperare 30.04.2018 Fiecare ProfesorVerinceanu AlexandruÎncă nu există evaluări
- 2019-20120 Orar Fiecare Clasa Since 16.09.2019Document38 pagini2019-20120 Orar Fiecare Clasa Since 16.09.2019Verinceanu AlexandruÎncă nu există evaluări
- 2019-20120 Orar Centralizator Clase Since 16.09.2019 SCOLIIDocument1 pagină2019-20120 Orar Centralizator Clase Since 16.09.2019 SCOLIIVerinceanu AlexandruÎncă nu există evaluări
- Formular 2 CERERE DE DESCHIDERE CONT PERSOANE FIZICE MINOREDocument4 paginiFormular 2 CERERE DE DESCHIDERE CONT PERSOANE FIZICE MINOREVerinceanu AlexandruÎncă nu există evaluări
- Formular 1 Cerere Contract DepozitDocument2 paginiFormular 1 Cerere Contract DepozitVerinceanu AlexandruÎncă nu există evaluări
- Orar Recuperare 30.04.2018 Fiecare ClasaDocument37 paginiOrar Recuperare 30.04.2018 Fiecare ClasaVerinceanu AlexandruÎncă nu există evaluări
- Adeverinta de SalariuDocument1 paginăAdeverinta de SalariuVerinceanu AlexandruÎncă nu există evaluări
- Formular 1 CERERE DE DESCHIDERE CONT PERSOANE FIZICEDocument3 paginiFormular 1 CERERE DE DESCHIDERE CONT PERSOANE FIZICEVerinceanu AlexandruÎncă nu există evaluări
- F1 - Cerere de Credit Persoane FiziceDocument16 paginiF1 - Cerere de Credit Persoane FiziceDaniel VichiluÎncă nu există evaluări
- Subiecte Virusuri Si Ciuperci BileteDocument30 paginiSubiecte Virusuri Si Ciuperci BiletecontpentruscoalaÎncă nu există evaluări
- Curs 5 Dinamica Raspunsului Imun CelularDocument4 paginiCurs 5 Dinamica Raspunsului Imun CelularAdaÎncă nu există evaluări