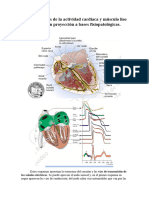
S-ar putea să vă placă și
- Anatomia y Fisiologia Del Sistema Excito ConductorDocument16 paginiAnatomia y Fisiologia Del Sistema Excito Conductorrosica1973Încă nu există evaluări
- Diploma E-Learning - Taquiarritmias SupraventricularesDocument73 paginiDiploma E-Learning - Taquiarritmias SupraventricularesFernando ArancibiaÎncă nu există evaluări
- Electrofisiología cardíaca: potenciales y conducción eléctricaDocument8 paginiElectrofisiología cardíaca: potenciales y conducción eléctricaAndrea Valeria Montenegro Reyes100% (1)
- CARDIO - Bases Biofísicas de La Actividad Cardiaca y Músculo Liso Vascular Con Proyección A Bases FisiopatológicasDocument28 paginiCARDIO - Bases Biofísicas de La Actividad Cardiaca y Músculo Liso Vascular Con Proyección A Bases FisiopatológicasRomina NogueiraÎncă nu există evaluări
- Génesis Del ECG NormalDocument8 paginiGénesis Del ECG NormalCamila CarpioÎncă nu există evaluări
- Electrofisiologia PDFDocument90 paginiElectrofisiologia PDFFabiola NorabuenaÎncă nu există evaluări
- Ecg CastellanosDocument143 paginiEcg CastellanosVelia Gallegos100% (1)
- Practica Potencial de Acción CardiacoDocument9 paginiPractica Potencial de Acción CardiacoXavi MGÎncă nu există evaluări
- Potenciales de acción y contracción del músculo cardíacoDocument8 paginiPotenciales de acción y contracción del músculo cardíacolilianaÎncă nu există evaluări
- Origen y Propagación 2Document9 paginiOrigen y Propagación 2Gabriela GregórioÎncă nu există evaluări
- ARRITMIASDocument21 paginiARRITMIASJoselyn RodriguezÎncă nu există evaluări
- Seminario Tp4 Fisica BiologicaDocument8 paginiSeminario Tp4 Fisica BiologicaEstefanía MansillaÎncă nu există evaluări
- Biofisica CardíacaDocument28 paginiBiofisica CardíacaMarcos Días100% (1)
- POTENCIAL DE MEMBRANA Y CARDIOVASCULADocument7 paginiPOTENCIAL DE MEMBRANA Y CARDIOVASCULAJacki ServianÎncă nu există evaluări
- 07 - Propiedades Del Músculo CardiacoDocument11 pagini07 - Propiedades Del Músculo CardiacoAlejandra HernandezÎncă nu există evaluări
- Corazón como bombaDocument7 paginiCorazón como bombaDante ValvassoriÎncă nu există evaluări
- Clase 3 4-10Document4 paginiClase 3 4-10Valentina UbillaÎncă nu există evaluări
- Fisiología cardíaca: estructura y función del corazónDocument67 paginiFisiología cardíaca: estructura y función del corazónMayreamar EstradaÎncă nu există evaluări
- Electrocardiograma Normal y Patológico - Medicina InternaDocument35 paginiElectrocardiograma Normal y Patológico - Medicina InternaPedro HermosoÎncă nu există evaluări
- SISTEMA CIRCULATORIODocument7 paginiSISTEMA CIRCULATORIOZahira AbrahamÎncă nu există evaluări
- Desfibrilador CardiacoDocument46 paginiDesfibrilador CardiacoJesus Resta NavarroÎncă nu există evaluări
- Ut II Guia10Document9 paginiUt II Guia10BarbaraAndradeQuirozÎncă nu există evaluări
- BiopotencialesDocument20 paginiBiopotencialesRoland CaixaÎncă nu există evaluări
- ECG Fisiología cardiovascularDocument22 paginiECG Fisiología cardiovascularKevin Argotti CazarÎncă nu există evaluări
- Mecanismos electrofisiológicos de la elevación del ST durante el infarto agudo de miocardioDocument7 paginiMecanismos electrofisiológicos de la elevación del ST durante el infarto agudo de miocardioElíseo E LQÎncă nu există evaluări
- Tarea 2 - Miguel Angel Ahumada Segura!!!Document16 paginiTarea 2 - Miguel Angel Ahumada Segura!!!MIGUEL ANGEL AHUMADAÎncă nu există evaluări
- Manual Toma e Interpretacion de ElectrocardiogramaDocument29 paginiManual Toma e Interpretacion de ElectrocardiogramaGuillermina SanchezÎncă nu există evaluări
- Aparato Cardiovascular 1T1Document41 paginiAparato Cardiovascular 1T1Marycielo LmÎncă nu există evaluări
- Fisiología - Guía # 1. Actividad Electrica Cardiaca 2023-2Document10 paginiFisiología - Guía # 1. Actividad Electrica Cardiaca 2023-2LAURA SOFIA PEREZ CUELLOÎncă nu există evaluări
- CARDIO!Document31 paginiCARDIO!Mauricio VianaÎncă nu există evaluări
- Propiedades Del Corazon de Sapo - CompressDocument23 paginiPropiedades Del Corazon de Sapo - CompressJUAN RELUZ ZAMBRANOÎncă nu există evaluări
- Electrofisiología CardíacaDocument13 paginiElectrofisiología CardíacajohanaÎncă nu există evaluări
- 1 EkgDocument12 pagini1 EkgegleeÎncă nu există evaluări
- Teoria Delm Musculo CardiacoDocument12 paginiTeoria Delm Musculo CardiacoAlbert KantÎncă nu există evaluări
- Marco Teorico ECGDocument11 paginiMarco Teorico ECGEliana Caceres TorricoÎncă nu există evaluări
- EKG Normal 1 y 2Document12 paginiEKG Normal 1 y 2Franck FarreraÎncă nu există evaluări
- GUÍA DE PRACTICA 3 Efecto de Los Iones Sobre La Frecuencia CardíacaDocument4 paginiGUÍA DE PRACTICA 3 Efecto de Los Iones Sobre La Frecuencia CardíacaArely Mical SánchezÎncă nu există evaluări
- Documento Sin TítuloDocument15 paginiDocumento Sin TítuloGuillermina SanchezÎncă nu există evaluări
- MielinizacionDocument57 paginiMielinizacionVictoria Castillo100% (1)
- Potencial de acción cardiaco y electrofisiologíaDocument14 paginiPotencial de acción cardiaco y electrofisiologíaluispin1186Încă nu există evaluări
- Electrofisiología y EcgDocument11 paginiElectrofisiología y Ecgivanlalvarez.22Încă nu există evaluări
- Automatismo AnormalDocument3 paginiAutomatismo AnormalEmily Janice Oliva AlburquequeÎncă nu există evaluări
- Propiedades eléctricas del corazónDocument4 paginiPropiedades eléctricas del corazónRichard Armijo GarridoÎncă nu există evaluări
- Guía de diapositivas sobre antiarrítmicosDocument11 paginiGuía de diapositivas sobre antiarrítmicosJoséÎncă nu există evaluări
- Electrofisiologia Cardiaca CMDM PDFDocument10 paginiElectrofisiologia Cardiaca CMDM PDFWendy OrtizÎncă nu există evaluări
- Biofísica Cardiaca - YGDocument3 paginiBiofísica Cardiaca - YGmichaellaÎncă nu există evaluări
- BiopotencialesDocument24 paginiBiopotencialesMiguel SoberanesÎncă nu există evaluări
- Electrocardiograma IDocument19 paginiElectrocardiograma IGeraldine LópezÎncă nu există evaluări
- Tarea 2 - Trabajo ColaborativoDocument46 paginiTarea 2 - Trabajo ColaborativoJuan Carlos Grisales MenesesÎncă nu există evaluări
- Cardiología CTODocument95 paginiCardiología CTOEfren Penaflores100% (1)
- UTII 1 Practica Potencial de Accion CardiacoDocument13 paginiUTII 1 Practica Potencial de Accion CardiacoMarisol RodríguezÎncă nu există evaluări
- Apunte EelctrocardiografiaDocument84 paginiApunte EelctrocardiografiaMelissa JerezÎncă nu există evaluări
- Teoría-Función Cardiovascular - MEHU 632Document24 paginiTeoría-Función Cardiovascular - MEHU 632Sara ParedesÎncă nu există evaluări
- Electrocardiograma 01Document15 paginiElectrocardiograma 01Yasirys Mariell PichardoÎncă nu există evaluări
- Cómo se comunican las neuronas: El milagro de la transmisión sinápticaDe la EverandCómo se comunican las neuronas: El milagro de la transmisión sinápticaÎncă nu există evaluări
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De la EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Evaluare: 5 din 5 stele5/5 (9)
- Medicina energética: Acupuntura 3 Circuitos energéticos secundariosDe la EverandMedicina energética: Acupuntura 3 Circuitos energéticos secundariosEvaluare: 5 din 5 stele5/5 (5)
- Temas de GinecologíaDocument10 paginiTemas de GinecologíaArlette AntoineÎncă nu există evaluări
- Ruptura Viscera Hueca y MacizaDocument29 paginiRuptura Viscera Hueca y MacizaJose Angel Loayza MozoÎncă nu există evaluări
- Equiipo de Intubacion Traqueal y BronquialDocument15 paginiEquiipo de Intubacion Traqueal y BronquialDavid González GamboaÎncă nu există evaluări
- Cardio 1Document9 paginiCardio 1Elizabeth RamírezÎncă nu există evaluări
- Protocolo de Anestesia en El Pre - Trans y Post Opera To RioDocument12 paginiProtocolo de Anestesia en El Pre - Trans y Post Opera To RioLibertadValenzuelaÎncă nu există evaluări
- Propiedades macroestructura maderaDocument11 paginiPropiedades macroestructura maderaCamilo Soruco SánchezÎncă nu există evaluări
- Los Productos OmnilifeDocument3 paginiLos Productos OmnilifeOscarin Veliz FelixÎncă nu există evaluări
- Evaluación sensorial de alimentosDocument34 paginiEvaluación sensorial de alimentosNaomi Hernandez GutiérrezÎncă nu există evaluări
- 12 Guia Practica de Comportamiento en Quirofano TSCDocument30 pagini12 Guia Practica de Comportamiento en Quirofano TSCSäyurîî PäntøjäÎncă nu există evaluări
- ArdsDocument38 paginiArdsKevin GarayÎncă nu există evaluări
- Sesión 5 Sistema EndocrinoDocument28 paginiSesión 5 Sistema Endocrinoanthony diaz100% (1)
- Observación de Células Animales y VegetalesDocument4 paginiObservación de Células Animales y VegetalescamilaÎncă nu există evaluări
- Manejo de Dolor Post OperatorioDocument37 paginiManejo de Dolor Post OperatorioMarcia Nely Escobar CastilloÎncă nu există evaluări
- 6.patrones Histológicos de Lesión GlomerularDocument13 pagini6.patrones Histológicos de Lesión Glomerularlhumbertocc100% (2)
- Adultez Intermedia-22Document10 paginiAdultez Intermedia-22Carlos Andres ArellanoÎncă nu există evaluări
- Diabetes BorradorDocument19 paginiDiabetes BorradorPriPorrasÎncă nu există evaluări
- Partes Internas RonaldoDocument15 paginiPartes Internas RonaldoJESSICA RUIZ ROLDANÎncă nu există evaluări
- Cuestionario de Fisiologia de La Reproducción y LactaciónDocument4 paginiCuestionario de Fisiologia de La Reproducción y LactaciónAndreaCrodaAndradeÎncă nu există evaluări
- Estructura Primaria y Secundaria Del TalloDocument44 paginiEstructura Primaria y Secundaria Del TalloCarlos Amezquita0% (1)
- Inmunologia Antigenos - AbbasDocument21 paginiInmunologia Antigenos - AbbasGarras BarriosÎncă nu există evaluări
- AnticoagulantesDocument24 paginiAnticoagulantesFernando MarquesÎncă nu există evaluări
- 0 Anhelando Su CompañeroDocument20 pagini0 Anhelando Su Compañerocamaguza28Încă nu există evaluări
- DOCUMENTODocument6 paginiDOCUMENTOLuis Angel Quiñones Saavedra100% (1)
- Procesos y Teorias Del EnvejecimientoDocument59 paginiProcesos y Teorias Del EnvejecimientoAlejandroÎncă nu există evaluări
- Lipogénesis PDFDocument19 paginiLipogénesis PDFfermin100% (1)
- Biofisica IDocument8 paginiBiofisica ILuxemburgoÎncă nu există evaluări
- Dieta CariogenicaDocument6 paginiDieta CariogenicaClaudia Lisset Baquedano BustamanteÎncă nu există evaluări
- Catalogo Laca Primavera DigitalDocument120 paginiCatalogo Laca Primavera DigitalBrenda MendezÎncă nu există evaluări
- Experimento - GlobosDocument7 paginiExperimento - GlobosluzÎncă nu există evaluări
- Escuela de EnfermeríaDocument46 paginiEscuela de EnfermeríaCarolina GuerreroÎncă nu există evaluări