S-ar putea să vă placă și
- EveryDay AyurvedaDocument11 paginiEveryDay AyurvedacvmraviÎncă nu există evaluări
- Professional Anesthesia HandbookDocument114 paginiProfessional Anesthesia HandbookSooryajith DineshÎncă nu există evaluări
- Drug Cards 1Document20 paginiDrug Cards 1Keying Chen100% (1)
- Sufixe Si PrefixeDocument14 paginiSufixe Si Prefixeseaofdreams6Încă nu există evaluări
- Normal ValuesDocument2 paginiNormal ValuesCheche Quirimit100% (1)
- BUN and CreatinineDocument4 paginiBUN and Creatininesarguss14100% (1)
- Viscerosomatic Reflexes: System/Organ Sympathetic ParasympatheticDocument2 paginiViscerosomatic Reflexes: System/Organ Sympathetic ParasympatheticParth PatelÎncă nu există evaluări
- Psiquiatria MolecularDocument426 paginiPsiquiatria Molecularfabriciobiblos100% (1)
- Assay Formats For Use R-Time PCRDocument14 paginiAssay Formats For Use R-Time PCRu77Încă nu există evaluări
- 10-Bad Moon WaningDocument22 pagini10-Bad Moon WaningAllen Zimmerman100% (1)
- Dengue Prevention and Control ProgramDocument7 paginiDengue Prevention and Control Programhazel mirandaÎncă nu există evaluări
- Trigeminal Nerve: Dr.B.B.GosaiDocument76 paginiTrigeminal Nerve: Dr.B.B.GosaiAlexAlxÎncă nu există evaluări
- Actual Issues of Environmental Economics: September 2018Document133 paginiActual Issues of Environmental Economics: September 2018Sharifa OraibieÎncă nu există evaluări
- Theobald Smith-The Discoverer of Ticks As Vectors of DiseaseDocument4 paginiTheobald Smith-The Discoverer of Ticks As Vectors of DiseaseJuan José Camacho T.Încă nu există evaluări
- AIP Asep Ema Santi RaihanDocument11 paginiAIP Asep Ema Santi RaihanAysha Siddika MimÎncă nu există evaluări
- Ernst Haeckel (1834-1919) : The German Darwin and His Impact On Modern BiologyDocument8 paginiErnst Haeckel (1834-1919) : The German Darwin and His Impact On Modern BiologyroraysideblancoÎncă nu există evaluări
- The Relationship Between Religiosity and Anxiety: A Meta-AnalysisDocument13 paginiThe Relationship Between Religiosity and Anxiety: A Meta-AnalysisRisdaFitriaÎncă nu există evaluări
- Monthly Topper Articles - International Journal of Science and Research (IJSR) DW WWDocument2 paginiMonthly Topper Articles - International Journal of Science and Research (IJSR) DW WWKomal KarÎncă nu există evaluări
- Gillon PDFDocument11 paginiGillon PDFEE KMÎncă nu există evaluări
- Web of Science Master Journal List - SearchDocument3 paginiWeb of Science Master Journal List - SearchAbdullahChaudhryÎncă nu există evaluări
- Introductionto ScienceDocument2 paginiIntroductionto ScienceMiguel SilvaÎncă nu există evaluări
- Psicologia AnomalísticaDocument6 paginiPsicologia AnomalísticaLuiza Fernandes BezerraÎncă nu există evaluări
- List of Life SciencesDocument1 paginăList of Life SciencesNathan Stuart The Retarded idiotÎncă nu există evaluări
- Introductionto Science 333333333333333335354545462Document2 paginiIntroductionto Science 333333333333333335354545462Ryan PointerÎncă nu există evaluări
- Introductionto ScienceDocument2 paginiIntroductionto ScienceJulhayda FernandoÎncă nu există evaluări
- Cooling Intervention Studies Among Outdoor Occupational Groups - A Review of The LiteratureDocument26 paginiCooling Intervention Studies Among Outdoor Occupational Groups - A Review of The Literatureวรศิษฐ์ อ๋องÎncă nu există evaluări
- Refine Your Search Results: Already Have A Manuscript?Document5 paginiRefine Your Search Results: Already Have A Manuscript?Knjižnica OŽB Požega PožegaÎncă nu există evaluări
- 1988 Dhondtcarryingcapacity Acta OeologiaDocument11 pagini1988 Dhondtcarryingcapacity Acta OeologiazunigagÎncă nu există evaluări
- Introductionto ScienceDocument2 paginiIntroductionto ScienceHarshini BaskaranÎncă nu există evaluări
- LiesdeAnatomia ParteIIDocument10 paginiLiesdeAnatomia ParteIIGleydson ResendeÎncă nu există evaluări
- Marx's Ecology. Materialism and Nature by John Bellamy FosterDocument4 paginiMarx's Ecology. Materialism and Nature by John Bellamy FosterSamara da Cunha OliveiraÎncă nu există evaluări
- Web of Science Master Journal List - SearchDocument7 paginiWeb of Science Master Journal List - Searchayoube mecaÎncă nu există evaluări
- Understanding Scientific Journals and Articles - Process of Science - VisionlearningDocument10 paginiUnderstanding Scientific Journals and Articles - Process of Science - VisionlearningnemarcosufvÎncă nu există evaluări
- Effects of Sunlight Exposure For Treatment of Tuberculosis: Literature ReviewDocument9 paginiEffects of Sunlight Exposure For Treatment of Tuberculosis: Literature ReviewFida NadhirÎncă nu există evaluări
- ArtyDocument3 paginiArtymsaidsaidyoussefÎncă nu există evaluări
- 2014.70.AntioxidantActivityofEndophytic Fungi Isolatedfrom LanneacoromendalicaDocument6 pagini2014.70.AntioxidantActivityofEndophytic Fungi Isolatedfrom LanneacoromendalicaBritish PropolisÎncă nu există evaluări
- Science in The News: BIOL1001 Lab Project Prepared by Dr. Charles Willis, Dr. Lorelei Patrick, Thomas FeltonDocument6 paginiScience in The News: BIOL1001 Lab Project Prepared by Dr. Charles Willis, Dr. Lorelei Patrick, Thomas FeltonArchie BrownÎncă nu există evaluări
- Review of Literature AyurvedaDocument9 paginiReview of Literature Ayurvedarrrrrr6650% (2)
- Case StudiesDocument3 paginiCase Studiesmaria ordonezÎncă nu există evaluări
- Ref 1. Allergy, Histamine and AntihistaminesDocument12 paginiRef 1. Allergy, Histamine and AntihistaminesCarolina DeaÎncă nu există evaluări
- CardenasCallirgos2019 ParasitosisPulmonaresDeMetastrongylusCutervoDocument3 paginiCardenasCallirgos2019 ParasitosisPulmonaresDeMetastrongylusCutervoAngelSaulSerranoAnguloÎncă nu există evaluări
- 3.6 - Industrial Ecology - Shedding More Light On Its Perspective of Understanding Nature As ModelDocument16 pagini3.6 - Industrial Ecology - Shedding More Light On Its Perspective of Understanding Nature As ModelKaroline Brito Coutinho FerreiraÎncă nu există evaluări
- Winging ItDocument4 paginiWinging ItFakename0% (2)
- Clinical Biochemistry of Domestic AnimalsDocument3 paginiClinical Biochemistry of Domestic AnimalsLuiz D'AngeloÎncă nu există evaluări
- Science:, 1691 (2003) Tehshik P. Yoon and Eric N. JacobsenDocument4 paginiScience:, 1691 (2003) Tehshik P. Yoon and Eric N. Jacobsen0191710017 JULIAN DAVID QUIMBAYO PARRA ESTUDIANTE ACTIVOÎncă nu există evaluări
- Sub 2 Present Knowledge HaemoDocument7 paginiSub 2 Present Knowledge HaemoOca DyahÎncă nu există evaluări
- Meneses 2018Document12 paginiMeneses 2018Heine MüllerÎncă nu există evaluări
- 121 AppraisalDocument14 pagini121 AppraisalKristine FloresÎncă nu există evaluări
- Oximetry-Guided Normoxic Resuscitation Following Canine Cardiac Arrest Reduces Cerebellar Purkinje Neuronal Damage - ResuscitationDocument2 paginiOximetry-Guided Normoxic Resuscitation Following Canine Cardiac Arrest Reduces Cerebellar Purkinje Neuronal Damage - ResuscitationCarlos GomesÎncă nu există evaluări
- Oxfordhb 9780198733522 Chapter 8-1Document17 paginiOxfordhb 9780198733522 Chapter 8-1Ahmed HusseinÎncă nu există evaluări
- Sample Poster For Lab 9Document1 paginăSample Poster For Lab 9tildalivÎncă nu există evaluări
- Anthropology of Food: January 2012Document9 paginiAnthropology of Food: January 2012alexacrystallinÎncă nu există evaluări
- Counting On Citations A Flawed Way To Measure QualityDocument3 paginiCounting On Citations A Flawed Way To Measure QualityLivia_BocalonÎncă nu există evaluări
- Writing A Better Research ArticleDocument13 paginiWriting A Better Research ArticleHelena GregórioÎncă nu există evaluări
- CapaCirurgiaGeral PDFDocument3 paginiCapaCirurgiaGeral PDFEmerson SilvaÎncă nu există evaluări
- Comparative Anatomyofthe VertebratesDocument21 paginiComparative Anatomyofthe VertebratesPatrick LentejasÎncă nu există evaluări
- Estimation of Bee Size Using Intertegular Span (Apoidea)Document4 paginiEstimation of Bee Size Using Intertegular Span (Apoidea)Robin Bedolla CochetÎncă nu există evaluări
- English Template (2022)Document4 paginiEnglish Template (2022)felix suhendraÎncă nu există evaluări
- Web of Science Coverage: General InformationDocument1 paginăWeb of Science Coverage: General InformationM. Hakkı YıldırımÎncă nu există evaluări
- Garlic As An Insect Repellent: JAMA The Journal of The American Medical Association September 2000Document4 paginiGarlic As An Insect Repellent: JAMA The Journal of The American Medical Association September 2000Eunice Kyla MapisaÎncă nu există evaluări
- La Resistencia Al Cambio en Psicoterapia: El Papel de La Reactancia, La Construcción Del Sí Mismo y El Tipo de DemandaDocument28 paginiLa Resistencia Al Cambio en Psicoterapia: El Papel de La Reactancia, La Construcción Del Sí Mismo y El Tipo de DemandaAbiÎncă nu există evaluări
- Competiția Interspecifică La Ciprinidele Din Ecosistemele Limnice Observații EtologiceDocument20 paginiCompetiția Interspecifică La Ciprinidele Din Ecosistemele Limnice Observații EtologiceMD1000Încă nu există evaluări
- E027308.full RecomendadaDocument10 paginiE027308.full RecomendadaTamires NunesÎncă nu există evaluări
- Article Analysis Science and TechnologyDocument13 paginiArticle Analysis Science and Technologyronald chumbaÎncă nu există evaluări
- Citationindexijdvl784511-7168846 015928Document8 paginiCitationindexijdvl784511-7168846 015928DEEPANCHAKKARAVARTHY KÎncă nu există evaluări
- A Filosofia Da Natureza de Aristóteles: June 2004Document14 paginiA Filosofia Da Natureza de Aristóteles: June 2004Jaime BilaÎncă nu există evaluări
- Citationindexijdvl784511-7168846 015928Document8 paginiCitationindexijdvl784511-7168846 015928segnumutraÎncă nu există evaluări
- Getting Ready To Publish-HGDocument31 paginiGetting Ready To Publish-HGnomiad81Încă nu există evaluări
- Abstract of in Silico Screening of Natural Compounds As PotentR4Document2 paginiAbstract of in Silico Screening of Natural Compounds As PotentR4Komal KarÎncă nu există evaluări
- Personalitate Borderline Cu Functionare Inalta - Abordare Complexa: Psihiatrica, Psihologica Si Psihodinamica (Studiu de Caz)Document12 paginiPersonalitate Borderline Cu Functionare Inalta - Abordare Complexa: Psihiatrica, Psihologica Si Psihodinamica (Studiu de Caz)IraIrochkaÎncă nu există evaluări
- Caloric Restriction: A Key to Understanding and Modulating AgingDe la EverandCaloric Restriction: A Key to Understanding and Modulating AgingÎncă nu există evaluări
- Methods For Studying CytotoxicityDocument2 paginiMethods For Studying Cytotoxicityu77Încă nu există evaluări
- Microbial Diversity by DGGEDocument2 paginiMicrobial Diversity by DGGEu77Încă nu există evaluări
- The Effects of Methanol, Ethanol, Propanol and Butanol On Bacterial Attachment To SurfacesDocument9 paginiThe Effects of Methanol, Ethanol, Propanol and Butanol On Bacterial Attachment To Surfacesu77Încă nu există evaluări
- Chelex DNA Preparation Protocol ForDocument2 paginiChelex DNA Preparation Protocol Foru77Încă nu există evaluări
- Chelex Protocol SLBDocument2 paginiChelex Protocol SLBu77Încă nu există evaluări
- Denaturing Gradient Gel Electrophoresis of β-ThalassemiaDocument2 paginiDenaturing Gradient Gel Electrophoresis of β-Thalassemiau77Încă nu există evaluări
- Dna Extraction Using Chelex ResinDocument7 paginiDna Extraction Using Chelex Resinu77Încă nu există evaluări
- Syber Green Quantitative-PCRDocument60 paginiSyber Green Quantitative-PCRu77Încă nu există evaluări
- Chelex 100-Instruction ManualDocument14 paginiChelex 100-Instruction Manualu77Încă nu există evaluări
- Utility of The Housekeeping Genes 18S rRNA, B-Actin-04Document8 paginiUtility of The Housekeeping Genes 18S rRNA, B-Actin-04u77Încă nu există evaluări
- Roche Primer DimerDocument4 paginiRoche Primer Dimeru77Încă nu există evaluări
- Chelating Resin - Product.informationDocument2 paginiChelating Resin - Product.informationu77Încă nu există evaluări
- Sybr Green PCR Master Mix and RT-PCR: ProtocolDocument64 paginiSybr Green PCR Master Mix and RT-PCR: Protocolu77Încă nu există evaluări
- Quantitative RT-PCR Protocol (Sybr Green I)Document8 paginiQuantitative RT-PCR Protocol (Sybr Green I)u77Încă nu există evaluări
- Real Quant PCRDocument48 paginiReal Quant PCRu77Încă nu există evaluări
- Real Time PCRDocument7 paginiReal Time PCRu77Încă nu există evaluări
- Real-Time Amplification On The Rotor-Gene: Overview of The Chemistries and OptimizationsDocument50 paginiReal-Time Amplification On The Rotor-Gene: Overview of The Chemistries and Optimizationsu77Încă nu există evaluări
- Real-Time PCRDocument5 paginiReal-Time PCRu77Încă nu există evaluări
- Block - 3 - 2014 Block 3 BookDocument197 paginiBlock - 3 - 2014 Block 3 BookhappyhappylandÎncă nu există evaluări
- LysosomesDocument9 paginiLysosomesapi-318784000Încă nu există evaluări
- Anatomy From The DoctorDocument208 paginiAnatomy From The DoctorSweet manÎncă nu există evaluări
- Cvs - 10 ConsolidatedDocument37 paginiCvs - 10 ConsolidatedezhilÎncă nu există evaluări
- Normal Anatomy, Histology, and Spontaneous Pathology of The Nasal Cavity of The Cynomolgus Monkey (Macaca Fascicularis)Document19 paginiNormal Anatomy, Histology, and Spontaneous Pathology of The Nasal Cavity of The Cynomolgus Monkey (Macaca Fascicularis)Dina SartikaÎncă nu există evaluări
- Integrementary System 2Document16 paginiIntegrementary System 2meyerscÎncă nu există evaluări
- 11 Must To Raise PigsDocument21 pagini11 Must To Raise PigsNshim TheogeneÎncă nu există evaluări
- MosquitosDocument2 paginiMosquitosapi-275246146Încă nu există evaluări
- Food Borne Diseases and Its PreventionDocument181 paginiFood Borne Diseases and Its Preventionapi-19916399Încă nu există evaluări
- Consensus Report: Periodontic-Endodontic Lesions: Group MembersDocument2 paginiConsensus Report: Periodontic-Endodontic Lesions: Group MembersAlex Grande CancapaÎncă nu există evaluări
- Beauty and Aging BookDocument217 paginiBeauty and Aging Bookasifmujteba2587100% (6)
- Head Injury: Presenters: Danny Farahana Supervised By: Dr. Nithya RamanathanDocument58 paginiHead Injury: Presenters: Danny Farahana Supervised By: Dr. Nithya RamanathanMuvenn KannanÎncă nu există evaluări
- Dengue Fever and PregnancyDocument6 paginiDengue Fever and PregnancyImanuel Far-FarÎncă nu există evaluări
- Respiratory and Circulatory SystemsDocument4 paginiRespiratory and Circulatory SystemsRommel BauzaÎncă nu există evaluări
- RRLDocument14 paginiRRLFreisanChenMandumotanÎncă nu există evaluări
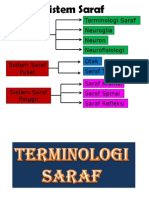
- Tisu Neural Neurofisiologi Neuron Neuroglia Terminologi SarafDocument141 paginiTisu Neural Neurofisiologi Neuron Neuroglia Terminologi SarafRainne LeeÎncă nu există evaluări
- Animal Testing Essential To Medical ProgressDocument7 paginiAnimal Testing Essential To Medical ProgressLayalÎncă nu există evaluări
- Kingdom AnimaliaDocument3 paginiKingdom AnimaliaLloydDagsil100% (1)
- ACT H Cracked Nipples FactSheet 2018-V1Document2 paginiACT H Cracked Nipples FactSheet 2018-V1dhea handyaraÎncă nu există evaluări
- B CellDocument10 paginiB CellSonia Elizabeth SimonÎncă nu există evaluări