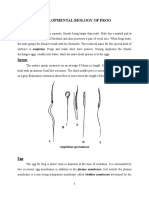
S-ar putea să vă placă și
- The Structure and Function of Animal Cell Components: An Introductory TextDe la EverandThe Structure and Function of Animal Cell Components: An Introductory TextÎncă nu există evaluări
- Spermatogenesis ReviewerDocument8 paginiSpermatogenesis ReviewerJowi Sal100% (1)
- Biotechnology and Food Safety: Proceedings of the Second International SymposiumDe la EverandBiotechnology and Food Safety: Proceedings of the Second International SymposiumDonald D. BillsEvaluare: 5 din 5 stele5/5 (1)
- L25 PDFDocument46 paginiL25 PDFMiles HuiÎncă nu există evaluări
- Kaufman’s Atlas of Mouse Development Supplement: With Coronal SectionsDe la EverandKaufman’s Atlas of Mouse Development Supplement: With Coronal SectionsRichard BaldockÎncă nu există evaluări
- Chick NeurulationDocument14 paginiChick NeurulationSophia RubiaÎncă nu există evaluări
- Neurulation Refers To The Folding Process in Vertebrate Embryos, Which Includes TheDocument6 paginiNeurulation Refers To The Folding Process in Vertebrate Embryos, Which Includes Theaaliya saaheenÎncă nu există evaluări
- Urogenital SystemDocument5 paginiUrogenital Systemstudent10100Încă nu există evaluări
- Respiratory System: StructureDocument29 paginiRespiratory System: StructureDr. Abir Ishtiaq100% (1)
- Spermatogenesis and OogenesisDocument23 paginiSpermatogenesis and OogenesisGerlJerl100% (1)
- SpermatogenesisDocument24 paginiSpermatogenesisOmm KoliÎncă nu există evaluări
- Early Embryonic DevelopmentDocument19 paginiEarly Embryonic DevelopmentderhangkerÎncă nu există evaluări
- 09 Exocytosis and EndocytosisDocument6 pagini09 Exocytosis and Endocytosisdrhendyjunior100% (1)
- Unit 13 Cleavage and GastrulationDocument26 paginiUnit 13 Cleavage and GastrulationFERHAEEZA KALAYAKANÎncă nu există evaluări
- Unit: 08 Placentation in Mammals: Developmental Biology Cr. Hours: 4 (3+1)Document30 paginiUnit: 08 Placentation in Mammals: Developmental Biology Cr. Hours: 4 (3+1)Task BirdÎncă nu există evaluări
- Introduction To Anatomy and PhysiologyDocument36 paginiIntroduction To Anatomy and PhysiologyKenneth Reigne ArguidasÎncă nu există evaluări
- Lab Report 4 GametogenesisDocument13 paginiLab Report 4 GametogenesisWnz NaiveÎncă nu există evaluări
- 01 - Embryology (Theory)Document54 pagini01 - Embryology (Theory)Simran KashyapÎncă nu există evaluări
- DJ (Microtubules, Microfilaments and Intermediate Filaments) - DJGFDocument43 paginiDJ (Microtubules, Microfilaments and Intermediate Filaments) - DJGFNick_989893Încă nu există evaluări
- MicrotubulesDocument39 paginiMicrotubulesAdnan QureshiÎncă nu există evaluări
- 07 Gastrulation BiologyDocument22 pagini07 Gastrulation BiologyKashif JamalÎncă nu există evaluări
- Laboratory Activity No. 9: The Frog Respiratory SystemDocument4 paginiLaboratory Activity No. 9: The Frog Respiratory SystemMaricris GuillermoÎncă nu există evaluări
- 01a Using Anatomical LanguageDocument6 pagini01a Using Anatomical Languageapi-3067625270% (1)
- Cilia and Flagella: Structure and Function in EukaryotesDocument10 paginiCilia and Flagella: Structure and Function in EukaryotesEr Purushottam PalÎncă nu există evaluări
- Activity 1: Frog's Egg, Cleavage, and Blastulation: Late-Yolk-Plug-4x-Ss - JPGDocument7 paginiActivity 1: Frog's Egg, Cleavage, and Blastulation: Late-Yolk-Plug-4x-Ss - JPGIvanÎncă nu există evaluări
- Intermediate FilamentsDocument4 paginiIntermediate FilamentsSai SridharÎncă nu există evaluări
- Exercise 1 The Cell 1Document6 paginiExercise 1 The Cell 1MJMadlangbayan100% (1)
- Syllabus v9 - Fall 2002 - Cell & Molecular BiologyDocument4 paginiSyllabus v9 - Fall 2002 - Cell & Molecular Biologyogangurel100% (2)
- 06 GastrulationtxtDocument38 pagini06 GastrulationtxtHafidzul HalimÎncă nu există evaluări
- Cell Structure and Its EnvironmentDocument50 paginiCell Structure and Its EnvironmentCarol FernandoÎncă nu există evaluări
- GastrulationDocument1 paginăGastrulationClaire BolalinÎncă nu există evaluări
- Chapter 28 The Reproductive SystemDocument40 paginiChapter 28 The Reproductive SystemRushaida AminullaÎncă nu există evaluări
- Cell Membrane (Structure and Function)Document150 paginiCell Membrane (Structure and Function)Dr. Tapan Kr. Dutta100% (1)
- Female Reproductive HistologyDocument59 paginiFemale Reproductive HistologyIta Indriani100% (2)
- 48 Hour ChickDocument13 pagini48 Hour ChickJowi SalÎncă nu există evaluări
- Cleavage: The Main Process of Animal DevelopmentDocument123 paginiCleavage: The Main Process of Animal DevelopmentNUR ASMA RIZKYÎncă nu există evaluări
- Cell Structures and Functions 02Document22 paginiCell Structures and Functions 02YASMEROÎncă nu există evaluări
- Physiology L6Document6 paginiPhysiology L6Anonymous elq7jZiSÎncă nu există evaluări
- Eye Chart 3 MTR A4 Paper PDFDocument1 paginăEye Chart 3 MTR A4 Paper PDFnaZarÎncă nu există evaluări
- Topic: Mechanisms of Fertilization:: Presented byDocument16 paginiTopic: Mechanisms of Fertilization:: Presented byMajani RajbonshiÎncă nu există evaluări
- Embryology - Updates and Highlights On Classic TopicsDocument222 paginiEmbryology - Updates and Highlights On Classic TopicsIndera Vyas100% (1)
- Lab Report Cell Bio MitosisDocument4 paginiLab Report Cell Bio Mitosisain syukriahÎncă nu există evaluări
- Lab 3 Diffusion and OsmosisDocument20 paginiLab 3 Diffusion and OsmosisrashadwcartwrightÎncă nu există evaluări
- A Brief History of BiologyDocument17 paginiA Brief History of BiologyCarlo AcebronÎncă nu există evaluări
- General Physiology: First YearDocument67 paginiGeneral Physiology: First YearGo HellÎncă nu există evaluări
- Histology of Glands: Dr. Zana TahseenDocument28 paginiHistology of Glands: Dr. Zana TahseenAhmed JawdetÎncă nu există evaluări
- Exercise 17 Serial Transverse Section of A 33 Hour Chick Embryo PDFDocument4 paginiExercise 17 Serial Transverse Section of A 33 Hour Chick Embryo PDFMichaela FayeÎncă nu există evaluări
- Early DevelopmentDocument5 paginiEarly DevelopmentMarina of The SeaÎncă nu există evaluări
- Notes On Cell OrganellesDocument11 paginiNotes On Cell Organellesapi-241062194Încă nu există evaluări
- Cell DifferentiationDocument16 paginiCell DifferentiationHarika KeshavÎncă nu există evaluări
- DOs and Donts in A BIOLABDocument7 paginiDOs and Donts in A BIOLABHaley BereyÎncă nu există evaluări
- Gametogenesis of Frog EmbryosDocument21 paginiGametogenesis of Frog EmbryosNics MartinezÎncă nu există evaluări
- Lymphoid SystemDocument11 paginiLymphoid SystemSheena PasionÎncă nu există evaluări
- Developmental BiologyDocument21 paginiDevelopmental BiologygahanÎncă nu există evaluări
- Embryonic Period: Dr. Khin Ma MADocument34 paginiEmbryonic Period: Dr. Khin Ma MAIbrahim Foondun100% (1)
- Cell Structure and Functions PPT 5Document24 paginiCell Structure and Functions PPT 5rajesh dua100% (1)
- Cell (Biochemical Aspects) : Dr. Abid AliDocument49 paginiCell (Biochemical Aspects) : Dr. Abid AliShahzad MazharÎncă nu există evaluări
- Developmental Biology of Frog: SpermDocument21 paginiDevelopmental Biology of Frog: SpermRavindra MadurÎncă nu există evaluări
- 2b. Types of Egg and CleavageDocument55 pagini2b. Types of Egg and CleavageManisha BishtÎncă nu există evaluări
- Digestive System HandoutsDocument7 paginiDigestive System HandoutsClexandrea Dela Luz Corpuz100% (1)
- NUS LSM4232 Lecture NotesDocument40 paginiNUS LSM4232 Lecture Notesijclement07Încă nu există evaluări
- Mammalian Oocyte Regulation Methoda and ProtocolesDocument316 paginiMammalian Oocyte Regulation Methoda and ProtocolesTlad AljazeraÎncă nu există evaluări
- Contoh PDFDocument9 paginiContoh PDFGratiano TelaumbanuaÎncă nu există evaluări
- CBSE Class 12 Biology Revision Notes Chapter-02 Sexual Reproduction in Flowering PlantsDocument11 paginiCBSE Class 12 Biology Revision Notes Chapter-02 Sexual Reproduction in Flowering PlantsNada NavasÎncă nu există evaluări
- Epigenetic Mating-Type InheritanceDocument17 paginiEpigenetic Mating-Type InheritancedanielÎncă nu există evaluări
- Quiz Microbial Genetics TortoraDocument17 paginiQuiz Microbial Genetics TortorapascabioÎncă nu există evaluări
- Plant Reproduction Guide STR FLOWER and POLINATIONDocument5 paginiPlant Reproduction Guide STR FLOWER and POLINATIONNicolas Schropp100% (1)
- Apoptosis: Protocols & Applications Guide Rev. 1/06Document23 paginiApoptosis: Protocols & Applications Guide Rev. 1/06Fidiya Septi Kusma WardaniÎncă nu există evaluări
- Replikasi, Transkripsi Dan Translasi DnaDocument19 paginiReplikasi, Transkripsi Dan Translasi DnaEllizabeth LilantiÎncă nu există evaluări
- Control of Gene ExpressionDocument20 paginiControl of Gene ExpressionBhaskar GangulyÎncă nu există evaluări
- pET-19b MapDocument2 paginipET-19b MapPaola BarbosaÎncă nu există evaluări
- CH 2 Rep. in Flowering Plants 2023 - 24 WSDocument3 paginiCH 2 Rep. in Flowering Plants 2023 - 24 WSArchfab73Încă nu există evaluări
- RNA InterferenceDocument23 paginiRNA Interferencechoryn modinaÎncă nu există evaluări
- Chapter 3 - Tissue Renewal, Regeneration, and RepairDocument12 paginiChapter 3 - Tissue Renewal, Regeneration, and RepairAgnieszka WisniewskaÎncă nu există evaluări
- Sexual Reproduction in Flowering PlantsDocument3 paginiSexual Reproduction in Flowering Plantssyed ayaaanÎncă nu există evaluări
- General Biology 1: Ms. Almira B. GacutanDocument27 paginiGeneral Biology 1: Ms. Almira B. GacutanBERNA MAE TAMAYOÎncă nu există evaluări
- Week 4 ExperimentDocument11 paginiWeek 4 ExperimentKomal KhanÎncă nu există evaluări
- ReproductionDocument162 paginiReproductionchumbavictor459100% (2)
- Study Guide Sexual Reproduction and Genetics English EditableDocument4 paginiStudy Guide Sexual Reproduction and Genetics English Editableapi-342334216Încă nu există evaluări
- Name: - Score: - Summative Test No. 1 in Science Second QuarterDocument19 paginiName: - Score: - Summative Test No. 1 in Science Second QuarterZyra Jean Castañeda Canseco100% (1)
- Introduction To Embryology 1Document72 paginiIntroduction To Embryology 1Sam MumoÎncă nu există evaluări
- Soalan 2Document19 paginiSoalan 2ROHAYU BINTI HARUN MoeÎncă nu există evaluări
- Genetics - NotesDocument3 paginiGenetics - NotesCristie Luz BautistaÎncă nu există evaluări
- CA Lesson 1 Cellular+ReproductionDocument60 paginiCA Lesson 1 Cellular+ReproductionNKAÎncă nu există evaluări
- ViewpdfDocument11 paginiViewpdfBhanu Pratap Singh TomarÎncă nu există evaluări
- Chapter 9 The Endocrine SystemDocument1 paginăChapter 9 The Endocrine SystemacÎncă nu există evaluări
- .Archivetempt3 CELL DIVISION - Q (14-18)Document5 pagini.Archivetempt3 CELL DIVISION - Q (14-18)Fatin HananiÎncă nu există evaluări
- ชีววิทยา สำหรับนักเรียนระดับมัธยมปลายDocument321 paginiชีววิทยา สำหรับนักเรียนระดับมัธยมปลายณัฐพล ศุภพิมลÎncă nu există evaluări
- Rafal CioskDocument3 paginiRafal CioskRigel_TÎncă nu există evaluări
- Genetics Revision Yr10Document10 paginiGenetics Revision Yr10Rachel QuahÎncă nu există evaluări
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDe la EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsEvaluare: 4.5 din 5 stele4.5/5 (6)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDe la EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisEvaluare: 3.5 din 5 stele3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDe la EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityEvaluare: 4 din 5 stele4/5 (3)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDe la EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceEvaluare: 4.5 din 5 stele4.5/5 (517)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeDe la EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeÎncă nu există evaluări
- Gut: the new and revised Sunday Times bestsellerDe la EverandGut: the new and revised Sunday Times bestsellerEvaluare: 4 din 5 stele4/5 (393)
- Tales from Both Sides of the Brain: A Life in NeuroscienceDe la EverandTales from Both Sides of the Brain: A Life in NeuroscienceEvaluare: 3 din 5 stele3/5 (18)
- Who's in Charge?: Free Will and the Science of the BrainDe la EverandWho's in Charge?: Free Will and the Science of the BrainEvaluare: 4 din 5 stele4/5 (65)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)De la EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Evaluare: 4 din 5 stele4/5 (411)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDe la Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessEvaluare: 4 din 5 stele4/5 (33)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperDe la EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperEvaluare: 4.5 din 5 stele4.5/5 (15)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesDe la EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesEvaluare: 4.5 din 5 stele4.5/5 (397)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedDe la EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedEvaluare: 4 din 5 stele4/5 (11)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionDe la EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionEvaluare: 4 din 5 stele4/5 (812)
- Good Without God: What a Billion Nonreligious People Do BelieveDe la EverandGood Without God: What a Billion Nonreligious People Do BelieveEvaluare: 4 din 5 stele4/5 (66)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorDe la EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorÎncă nu există evaluări
- Seven and a Half Lessons About the BrainDe la EverandSeven and a Half Lessons About the BrainEvaluare: 4 din 5 stele4/5 (109)
- Human: The Science Behind What Makes Your Brain UniqueDe la EverandHuman: The Science Behind What Makes Your Brain UniqueEvaluare: 3.5 din 5 stele3.5/5 (38)
- Lymph & Longevity: The Untapped Secret to HealthDe la EverandLymph & Longevity: The Untapped Secret to HealthEvaluare: 4.5 din 5 stele4.5/5 (13)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldDe la EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldEvaluare: 4 din 5 stele4/5 (595)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouDe la EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouEvaluare: 4.5 din 5 stele4.5/5 (62)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindDe la EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindEvaluare: 4.5 din 5 stele4.5/5 (93)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomDe la EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomEvaluare: 4 din 5 stele4/5 (216)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldDe la EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldEvaluare: 4.5 din 5 stele4.5/5 (18)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemDe la EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemEvaluare: 4.5 din 5 stele4.5/5 (115)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondDe la EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondEvaluare: 4 din 5 stele4/5 (4)