S-ar putea să vă placă și
- Programa PC 1Document37 paginiPrograma PC 1Carlos Fernando Soto AlonsoÎncă nu există evaluări
- Embarazo Normal - Modificaciones Gravidicas SistemicasDocument46 paginiEmbarazo Normal - Modificaciones Gravidicas SistemicasPaola GallardoÎncă nu există evaluări
- Isoinmunización Materno-FetalDocument24 paginiIsoinmunización Materno-FetalJorge KahvehÎncă nu există evaluări
- Ventiladores de Presion y VolumenDocument45 paginiVentiladores de Presion y VolumenSantiago Cruz MontesinosÎncă nu există evaluări
- Taller 5 CostoDocument7 paginiTaller 5 Costocristian alberto sierra maciasÎncă nu există evaluări
- Pae SarampionDocument25 paginiPae Sarampiondaniel gomez menesesÎncă nu există evaluări
- Protocolo de Problema de Desnutricion en NicaraguaDocument29 paginiProtocolo de Problema de Desnutricion en NicaraguaJuan Ponce VanegaÎncă nu există evaluări
- Fisiología Del EmbarazoDocument13 paginiFisiología Del EmbarazoKaren RivasplataÎncă nu există evaluări
- Inducción Del PartoDocument22 paginiInducción Del PartoJuan A. LópezÎncă nu există evaluări
- Signos de Alarma Recien NacidoDocument21 paginiSignos de Alarma Recien NacidoDiana Collado candelaÎncă nu există evaluări
- Patrones de Ocurrencia de EnfermedadesDocument17 paginiPatrones de Ocurrencia de EnfermedadesJesus Rivera100% (1)
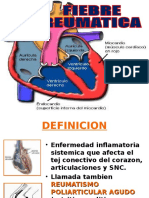
- Fiebre Reumatica Abril 2014Document16 paginiFiebre Reumatica Abril 2014Cindy Fiorella Aguado OrellanaÎncă nu există evaluări
- Trastornos Hipertensivos en El EmbarazoDocument24 paginiTrastornos Hipertensivos en El EmbarazomaycolupchbÎncă nu există evaluări
- Necesidad de HidratacionDocument3 paginiNecesidad de HidratacionStefany GonzalesÎncă nu există evaluări
- Caso Atención Prenatal ReenfocadaDocument3 paginiCaso Atención Prenatal ReenfocadaMilagros DDíazÎncă nu există evaluări
- Farmaco - Unidad 4 - Tema 28 - Farmacologia Inhibidores Uterinos - 09jul14Document22 paginiFarmaco - Unidad 4 - Tema 28 - Farmacologia Inhibidores Uterinos - 09jul14Ramsdry232Încă nu există evaluări
- Clase 10 - Familia - Enterob E. Coli, Sal, Shig, CÁCEDA 10Document130 paginiClase 10 - Familia - Enterob E. Coli, Sal, Shig, CÁCEDA 10lor bellidoÎncă nu există evaluări
- HOSPITAL BACA ORTIZ Exp. DeshidratacionDocument52 paginiHOSPITAL BACA ORTIZ Exp. Deshidratacionjavier patricio cando casanovaÎncă nu există evaluări
- Hemorragia Post PartoDocument19 paginiHemorragia Post PartoMarylin Quispe COrdova100% (1)
- Examenes de Lab Oratorio de QS, BH, TP, TTP, EGODocument6 paginiExamenes de Lab Oratorio de QS, BH, TP, TTP, EGOHaziel Venegas JimenezÎncă nu există evaluări
- 5 Estenosis PiloroDocument6 pagini5 Estenosis PiloroSheyla Ascencio OkonÎncă nu există evaluări
- Diagnóstico de Salud FetalDocument29 paginiDiagnóstico de Salud FetalJoSe Luis Quiroz Zela100% (1)
- Anestesia Pediatrica de Pacientescon Patologia Renal y EndocrinaDocument46 paginiAnestesia Pediatrica de Pacientescon Patologia Renal y EndocrinaDiego ManriqueÎncă nu există evaluări
- 04 Gastritis Aguda CronicaDocument58 pagini04 Gastritis Aguda CronicaAlex Trinidad Armas0% (1)
- Trabajo de Parto: Caso ClínicoDocument35 paginiTrabajo de Parto: Caso ClínicoMarcos BonillaÎncă nu există evaluări
- Taller T-StudentDocument2 paginiTaller T-StudentAbraham CorreaÎncă nu există evaluări
- Fisiopatología de La EspectoraciónDocument13 paginiFisiopatología de La EspectoraciónOskar ToscanoÎncă nu există evaluări
- GastroenteritisDocument33 paginiGastroenteritisMa. Laura CazarezÎncă nu există evaluări
- Salud Fetal Dra AlvarezDocument101 paginiSalud Fetal Dra AlvarezFernanda Catalina GamarraÎncă nu există evaluări
- Ictericia X Lactancia MDocument6 paginiIctericia X Lactancia MAnaly Cajusol DiazÎncă nu există evaluări
- Sindrome CarcinoideDocument31 paginiSindrome Carcinoideemely_eceÎncă nu există evaluări
- Hipoglicemia Neonatal JSabaDocument51 paginiHipoglicemia Neonatal JSabaAlejandra Medina0% (1)
- Practica #03..............Document26 paginiPractica #03..............Mejia Vilchez Lennan80% (5)
- Manejo de La Deshidratación en Pediatría, Resumen ClaseDocument6 paginiManejo de La Deshidratación en Pediatría, Resumen ClaseCarlos Danilo Noroña CÎncă nu există evaluări
- Hemorragia de La Primera Mitad Del EmbarazoDocument48 paginiHemorragia de La Primera Mitad Del EmbarazoamiraÎncă nu există evaluări
- Ejercicio BheDocument5 paginiEjercicio BheAngela PerisÎncă nu există evaluări
- Arbol Problemas SANDocument3 paginiArbol Problemas SANFabrizio GarciaÎncă nu există evaluări
- Guia de Liquidos y Electrolitos 2021-IIDocument6 paginiGuia de Liquidos y Electrolitos 2021-IICesar Augusto Fernandez BuenañoÎncă nu există evaluări
- Manejo Del Desnutrido Agudo SeveroDocument9 paginiManejo Del Desnutrido Agudo SeveroAlexisÎncă nu există evaluări
- HtaieDocument44 paginiHtaieJorge Luis Babilon AlibertoÎncă nu există evaluări
- Cambios Fisiologicos y Anatomicos en El EmbarazoDocument14 paginiCambios Fisiologicos y Anatomicos en El EmbarazoCrsithiamÎncă nu există evaluări
- Bienestar Fetal Insatisfactorio PDFDocument61 paginiBienestar Fetal Insatisfactorio PDFDiego Fernando Hoyos SÎncă nu există evaluări
- Gestante A Término Con Anemia Ferropenica y Bajo Peso Del Recién NacidoDocument26 paginiGestante A Término Con Anemia Ferropenica y Bajo Peso Del Recién NacidoSeleny Soto BenitesÎncă nu există evaluări
- SojaDocument15 paginiSojaDiana Gutierrez ChÎncă nu există evaluări
- RCTGDocument86 paginiRCTGPanda CastañedaÎncă nu există evaluări
- Actualizacion Rotura+Prematura+de+Membranas 2012 SEGODocument21 paginiActualizacion Rotura+Prematura+de+Membranas 2012 SEGOebama20Încă nu există evaluări
- Monitoreo FetalDocument6 paginiMonitoreo FetalWillan Garcia OrtizÎncă nu există evaluări
- Tratamiento de La Enfermedad Diarreica Según ElDocument19 paginiTratamiento de La Enfermedad Diarreica Según ElDiana CBÎncă nu există evaluări
- PH FecalDocument3 paginiPH FecalrellenodechileconcarÎncă nu există evaluări
- Caso Clinico de Hipoglicemia NeonatalDocument17 paginiCaso Clinico de Hipoglicemia NeonatalFaniizhiita AqÎncă nu există evaluări
- Pruebas Hematologicas en Palomas Castilla ' Columba LiviaDocument30 paginiPruebas Hematologicas en Palomas Castilla ' Columba LiviaErck Martin Alvarado NancayÎncă nu există evaluări
- Areas de Quirófano y Equipo MultidisciplinarioDocument9 paginiAreas de Quirófano y Equipo MultidisciplinarioALEJANDRA MENDEZÎncă nu există evaluări
- Formula para Calcular Superficie CorporalDocument1 paginăFormula para Calcular Superficie CorporalalexÎncă nu există evaluări
- Trabajo de Parto y Puerperio NormalDocument19 paginiTrabajo de Parto y Puerperio NormalAleja SalazarÎncă nu există evaluări
- Hongos en La Industria AlimentariaDocument3 paginiHongos en La Industria Alimentariaelson100% (1)
- Resveratrol en Enfermedades Del CorazónDocument32 paginiResveratrol en Enfermedades Del CorazónMiguelChavezÎncă nu există evaluări
- Hiperemesis GravidicaDocument23 paginiHiperemesis GravidicaHellenxita RiveraÎncă nu există evaluări
- Balance HidroelectroliticoDocument11 paginiBalance HidroelectroliticoGuadalupe Heredia50% (4)
- EcopatologíaDocument17 paginiEcopatologíaMarikitin CedeñoÎncă nu există evaluări
- Adaptacion de Tilapia A Agua SaladaDocument23 paginiAdaptacion de Tilapia A Agua SaladaMaria Carolina100% (2)
- Practica 6 FisioDocument7 paginiPractica 6 FisioAlejandro H Fisher50% (2)
- OPuOPr-informe S&H EngulianDocument4 paginiOPuOPr-informe S&H EngulianDamianFrancucciÎncă nu există evaluări
- Consecuencias Del Deterioro AmbientalDocument20 paginiConsecuencias Del Deterioro AmbientalJORGE RIVASÎncă nu există evaluări
- Endoscopia de ColumnaDocument31 paginiEndoscopia de ColumnaTOMASNOEL5100% (2)
- 17 Aviso de Accidente de Trabajo IESSDocument3 pagini17 Aviso de Accidente de Trabajo IESSJamil Valencia IntriagoÎncă nu există evaluări
- Mapa Conceptual Mamiferos.1Document1 paginăMapa Conceptual Mamiferos.1Ale Camargo67% (9)
- Requisitos para Abrir Una GasolineraDocument19 paginiRequisitos para Abrir Una GasolineraElmer Jesus Flores Mamani100% (2)
- Ejercicio 1 Prueba Sin CargaDocument2 paginiEjercicio 1 Prueba Sin CargaCarlos LopezÎncă nu există evaluări
- Ensayo de FilosofiaDocument5 paginiEnsayo de Filosofiamonica leichtleÎncă nu există evaluări
- Diseño de CavidadesDocument10 paginiDiseño de CavidadesCarlos Enrique Cuevas SuárezÎncă nu există evaluări
- Ficha de ReligiónDocument3 paginiFicha de ReligiónJavier CrisÎncă nu există evaluări
- Estimulacion Multisensorial Fisio PediatricoDocument11 paginiEstimulacion Multisensorial Fisio PediatricoJenny Andrea Rodriguez Quintero100% (1)
- Preguntas Practicas Tema 2Document59 paginiPreguntas Practicas Tema 2Saioa ArranzÎncă nu există evaluări
- Presentacion de Juma de Mexico 2020Document11 paginiPresentacion de Juma de Mexico 2020erickaÎncă nu există evaluări
- Normativa TFG 23-24 - 091023Document15 paginiNormativa TFG 23-24 - 091023angelalcazar1504Încă nu există evaluări
- Aifc Tarea Docente Semana 1 Erickson MoralesDocument17 paginiAifc Tarea Docente Semana 1 Erickson MoralesEricksonÎncă nu există evaluări
- Cuenca Hidrografica 2018-IIDocument58 paginiCuenca Hidrografica 2018-IIanon_487033747Încă nu există evaluări
- G4E HolisticSecurityGuide SpanishDocument20 paginiG4E HolisticSecurityGuide SpanishJahir CastroÎncă nu există evaluări
- Teratología, Agentes FetotóxicosDocument16 paginiTeratología, Agentes FetotóxicosJosé Luis Majano MurilloÎncă nu există evaluări
- Condensador ElectroliticoDocument7 paginiCondensador ElectroliticoAndres Felipe CarvajalÎncă nu există evaluări
- Proyecto Gerencia PDFDocument38 paginiProyecto Gerencia PDFjoseÎncă nu există evaluări
- Plan de Estudios KinesiologiaDocument11 paginiPlan de Estudios KinesiologiaDamii DinardiÎncă nu există evaluări
- Receta de PostreDocument25 paginiReceta de PostreYulianna EQ100% (1)
- Diarrea Aguda Infecciosa en Pediatría (DAI)Document42 paginiDiarrea Aguda Infecciosa en Pediatría (DAI)Gary Mártin Sandoval MerinoÎncă nu există evaluări
- PRÁCTICA 1.2 Parte-Ignacio Martínez y Livis RuizDocument4 paginiPRÁCTICA 1.2 Parte-Ignacio Martínez y Livis RuiznachoÎncă nu există evaluări
- HT 5227XX GUARD TANK STRUCTURAL ALUMINIUM (EID-187 V.02) 1.pdf FICHA TECNICADocument2 paginiHT 5227XX GUARD TANK STRUCTURAL ALUMINIUM (EID-187 V.02) 1.pdf FICHA TECNICADennis MendozaÎncă nu există evaluări
- Trastornos Hemodinamicos CR TestDocument207 paginiTrastornos Hemodinamicos CR TestJose Benito Guevara Conrado100% (1)
- NUTRICION Clase 4 TerminadoDocument7 paginiNUTRICION Clase 4 TerminadoANMI Al FinÎncă nu există evaluări
- Enfermedades Transmisibles y No TransmisiblesDocument7 paginiEnfermedades Transmisibles y No Transmisiblesnohelizdyana100% (1)