S-ar putea să vă placă și
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- Natural Bodybuilding Competition Preparation and Recovery - A 12-Month Case StudyDocument11 paginiNatural Bodybuilding Competition Preparation and Recovery - A 12-Month Case StudyRodrigo Castillo100% (1)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- A Review of The Carbohydrate-Insulin Model of ObesityDocument4 paginiA Review of The Carbohydrate-Insulin Model of ObesityAnonymous ZSmSZErTqtÎncă nu există evaluări
- Nutrition For Throwers, Jumpers and Combined Events AthletesDocument16 paginiNutrition For Throwers, Jumpers and Combined Events AthletesRodrigo CastilloÎncă nu există evaluări
- 2011 Compendium Tracking Guide MET INTENSITIES PDFDocument25 pagini2011 Compendium Tracking Guide MET INTENSITIES PDF1972197219721972Încă nu există evaluări
- Glycemic Index - Research & Nutrition Recommendations - 80 PagesDocument84 paginiGlycemic Index - Research & Nutrition Recommendations - 80 PageslozzzzzÎncă nu există evaluări
- Fluid Needs For Training and Competition in Athletics.Document15 paginiFluid Needs For Training and Competition in Athletics.Rodrigo CastilloÎncă nu există evaluări
- The Female Athlete Triad - Components, Nutrition Issues, and Health ConsequencesDocument17 paginiThe Female Athlete Triad - Components, Nutrition Issues, and Health ConsequencesRodrigo CastilloÎncă nu există evaluări
- Nutrition For The Young AthleteDocument18 paginiNutrition For The Young AthleteRodrigo CastilloÎncă nu există evaluări
- Innovations in Athletic Preparation - Role of Substrate Availability To Modify Training Adaptation and PerformanceDocument17 paginiInnovations in Athletic Preparation - Role of Substrate Availability To Modify Training Adaptation and PerformanceRodrigo CastilloÎncă nu există evaluări
- Sleeping Metabolic Rate in Relation To Body Mass Index and Body CompositionDocument8 paginiSleeping Metabolic Rate in Relation To Body Mass Index and Body CompositionRodrigo CastilloÎncă nu există evaluări
- Analysis of Energy Metabolism in Humans - A Review of MethodologiesDocument15 paginiAnalysis of Energy Metabolism in Humans - A Review of MethodologiesRodrigo CastilloÎncă nu există evaluări
- The mTOR Pathway - Implications For DNA ReplicationDocument9 paginiThe mTOR Pathway - Implications For DNA ReplicationRodrigo CastilloÎncă nu există evaluări
- Indirect Calorimetry - An Indispensable Tool To Understand and Predict ObesityDocument5 paginiIndirect Calorimetry - An Indispensable Tool To Understand and Predict ObesityRodrigo CastilloÎncă nu există evaluări
- "Calories In, Calories Out" and Macronutrient Intake: The Hope, Hype, and 1 Science of Calories.Document12 pagini"Calories In, Calories Out" and Macronutrient Intake: The Hope, Hype, and 1 Science of Calories.Nanci GuestÎncă nu există evaluări
- Calculation of Substrate Oxidation Rates in Vivo From Gaseous ExchangeDocument7 paginiCalculation of Substrate Oxidation Rates in Vivo From Gaseous ExchangeRodrigo CastilloÎncă nu există evaluări
- PGC-1 - The Energetic Regulator in Cardiac MetabolismDocument18 paginiPGC-1 - The Energetic Regulator in Cardiac MetabolismRodrigo CastilloÎncă nu există evaluări
- Nutrients 11 02171 v2Document9 paginiNutrients 11 02171 v2Rodrigo CastilloÎncă nu există evaluări
- Pi Is 0092867417301824Document17 paginiPi Is 0092867417301824bossanovasÎncă nu există evaluări
- International Journal of Sport Nutrition and Exercise Metabolism. This Article Appears Here in Its Accepted, PeerDocument21 paginiInternational Journal of Sport Nutrition and Exercise Metabolism. This Article Appears Here in Its Accepted, PeerRodrigo CastilloÎncă nu există evaluări
- 10 3945@ajcn 117 152777Document10 pagini10 3945@ajcn 117 152777Rodrigo CastilloÎncă nu există evaluări
- Gastric Emptying, Gastric Secretion and Enterogastrone Response After Administration of Milk Proteins or Their Peptide Hydrolysates in HumansDocument11 paginiGastric Emptying, Gastric Secretion and Enterogastrone Response After Administration of Milk Proteins or Their Peptide Hydrolysates in HumansRodrigo CastilloÎncă nu există evaluări
- Case Study - Natural Bodybuilding Contest PreparationDocument8 paginiCase Study - Natural Bodybuilding Contest PreparationRodrigo CastilloÎncă nu există evaluări
- Martins 2019Document38 paginiMartins 2019Rodrigo CastilloÎncă nu există evaluări
- Validation of An Integrated Pedal Desk and Electronic Behavior Tracking PlatformDocument9 paginiValidation of An Integrated Pedal Desk and Electronic Behavior Tracking PlatformRodrigo CastilloÎncă nu există evaluări
- CADocument9 paginiCARodrigo CastilloÎncă nu există evaluări
- Journal of Sport Scinece SPECIAL ISSUE IAAF NUTRITION SUPPLEMENTDocument1 paginăJournal of Sport Scinece SPECIAL ISSUE IAAF NUTRITION SUPPLEMENTRodrigo CastilloÎncă nu există evaluări
- Gastric Emptying, Gastric Secretion and Enterogastrone Response After Administration of Milk Proteins or Their Peptide Hydrolysates in HumansDocument11 paginiGastric Emptying, Gastric Secretion and Enterogastrone Response After Administration of Milk Proteins or Their Peptide Hydrolysates in HumansRodrigo CastilloÎncă nu există evaluări
- Business Model Canvas TemplateDocument3 paginiBusiness Model Canvas TemplatehanadracoÎncă nu există evaluări
- Baar K 2006Document6 paginiBaar K 2006Rodrigo CastilloÎncă nu există evaluări
- Carbohydrates and Fat For Training and RecoveryDocument16 paginiCarbohydrates and Fat For Training and RecoveryRodrigo CastilloÎncă nu există evaluări
- Wendler ConditioningDocument1 paginăWendler ConditioningMatthew CittaÎncă nu există evaluări
- Types of Muscles: Muscular SystemDocument13 paginiTypes of Muscles: Muscular SystemHaru MatsubuchiÎncă nu există evaluări
- Tétel SportDocument6 paginiTétel SportRomina NyersÎncă nu există evaluări
- Knes 387 Combatives Lesson PlanDocument9 paginiKnes 387 Combatives Lesson Planapi-645072592Încă nu există evaluări
- Time Course of Recovery Following Resistance Training LeadingDocument13 paginiTime Course of Recovery Following Resistance Training LeadingPablo PadrónÎncă nu există evaluări
- Encountering T. KrishnamacharyaDocument6 paginiEncountering T. KrishnamacharyaAnthonyÎncă nu există evaluări
- Discover Proprioception: A "Hidden" Sense©: by Zoe Mailloux, OTD, OTR/L, FAOTADocument2 paginiDiscover Proprioception: A "Hidden" Sense©: by Zoe Mailloux, OTD, OTR/L, FAOTAFlahiza MarquesÎncă nu există evaluări
- 35Document3 pagini35Einah EinahÎncă nu există evaluări
- DA Form 705: APFT CardDocument6 paginiDA Form 705: APFT CardAllArmyRegulationsÎncă nu există evaluări
- Exercise Handouts Foam RollerDocument2 paginiExercise Handouts Foam RollerEd WeardÎncă nu există evaluări
- School of Mount St. Mary, Inc.: Flexible Instruction Delivery PlanDocument14 paginiSchool of Mount St. Mary, Inc.: Flexible Instruction Delivery PlanJaysonMananquilLabsan100% (1)
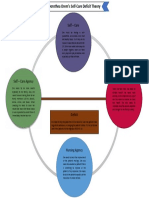
- Dorothea Orem's Self-Care Deficit TheoryDocument1 paginăDorothea Orem's Self-Care Deficit TheoryWerlyn Mae GonzagaÎncă nu există evaluări
- FREE January Kickstart With Rebecca LouiseDocument13 paginiFREE January Kickstart With Rebecca LouisenunimaliukÎncă nu există evaluări
- Strong HandsDocument6 paginiStrong HandsGábor KissÎncă nu există evaluări
- English PortfolioDocument8 paginiEnglish Portfoliodreamy-girlÎncă nu există evaluări
- Grade 12 Q1-Pe 3 Quaterly ExamDocument4 paginiGrade 12 Q1-Pe 3 Quaterly ExamRebecca Pidlaoan100% (1)
- Health Optimizing Physical Education 1: Jethro Leo P. Almaden Grade 11 - DuhatDocument22 paginiHealth Optimizing Physical Education 1: Jethro Leo P. Almaden Grade 11 - DuhatJethro Leo AlmadenÎncă nu există evaluări
- 9 Best TMJ Exercises For Pain ReliefDocument4 pagini9 Best TMJ Exercises For Pain ReliefAk KjÎncă nu există evaluări
- Anaerobixx Diet UPDATEDocument8 paginiAnaerobixx Diet UPDATEIlijaÎncă nu există evaluări
- Jepoy Swimming - Introduction and OrientationDocument37 paginiJepoy Swimming - Introduction and OrientationAce PortgasÎncă nu există evaluări
- Lesson Five: Vocabulary: Vocabulary WordsDocument3 paginiLesson Five: Vocabulary: Vocabulary WordsKristine BocciaÎncă nu există evaluări
- Giáo Trình LPTD 2Document40 paginiGiáo Trình LPTD 2Hưng Trịnh TrọngÎncă nu există evaluări
- FIN268 Lecture Notes 4Document5 paginiFIN268 Lecture Notes 4Joseph SibiÎncă nu există evaluări
- Om Yoga Magazine - January 2020Document134 paginiOm Yoga Magazine - January 2020dacostag0% (1)
- L1 - ACTIVITY 1 - PE4 - Renren Reyes IDocument2 paginiL1 - ACTIVITY 1 - PE4 - Renren Reyes IAira Y. PaglinawanÎncă nu există evaluări
- 21 Exercises For Injury Free Mass - T NationDocument18 pagini21 Exercises For Injury Free Mass - T Nationrfa940Încă nu există evaluări
- Pe Week 1 and 2Document36 paginiPe Week 1 and 2Czarina GenteroyÎncă nu există evaluări
- Healthy LifestyleDocument13 paginiHealthy Lifestylecoco jaiÎncă nu există evaluări
- Energy Manipulation Part IIDocument22 paginiEnergy Manipulation Part IIgrobi69100% (1)
- Oxygen DebtDocument3 paginiOxygen DebtrajeevkeyarÎncă nu există evaluări