S-ar putea să vă placă și
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (120)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Classification of Class Insecta Upto Orders PDFDocument3 paginiClassification of Class Insecta Upto Orders PDFAyush Sahu100% (1)
- Genetic Manipulation of Host PlantDocument15 paginiGenetic Manipulation of Host PlantDeza Grace Lobigas GerelingoÎncă nu există evaluări
- NeemAzal Leaflet WebDocument6 paginiNeemAzal Leaflet WebGyorgy SzigyartoÎncă nu există evaluări
- Georges Lakhovsky - The Secret of Life PDFDocument67 paginiGeorges Lakhovsky - The Secret of Life PDFAnonymous DqeRReÎncă nu există evaluări
- De TS Tieng Anh 10 Binh Duong 23 24Document6 paginiDe TS Tieng Anh 10 Binh Duong 23 24nhomly10a72023Încă nu există evaluări
- Diku Jackfruit (Recovered)Document24 paginiDiku Jackfruit (Recovered)purbaliÎncă nu există evaluări
- Sip (Science) (Insecticide)Document9 paginiSip (Science) (Insecticide)GabrielMichaelMalubayCapuyanÎncă nu există evaluări
- PSLE Science 2012 Booklet BDocument20 paginiPSLE Science 2012 Booklet BVenetia TohÎncă nu există evaluări
- Traitors TomeDocument50 paginiTraitors TomeRyan SikderÎncă nu există evaluări
- Arcadia Reptile 2018Document32 paginiArcadia Reptile 2018marianpcbÎncă nu există evaluări
- The Gut Microbiota of Insects - Diversity in Structure and FunctionDocument37 paginiThe Gut Microbiota of Insects - Diversity in Structure and FunctionribozymesÎncă nu există evaluări
- General KnowledgeDocument20 paginiGeneral KnowledgeAmanpreet KaurÎncă nu există evaluări
- Lesson Plan 1 GRADE 11Document4 paginiLesson Plan 1 GRADE 11Charmaine Cimanes BeaÎncă nu există evaluări
- Incect AnatomiaDocument14 paginiIncect AnatomiaKrisztina Margit HorváthÎncă nu există evaluări
- Morphogenetic Field Effects and ApplicationsDocument8 paginiMorphogenetic Field Effects and ApplicationsK-PAX100% (6)
- Body Structures of Animals For Survival - GR4Document30 paginiBody Structures of Animals For Survival - GR4Rjay CadaÎncă nu există evaluări
- Insect Life Cycle: A Reading A-Z Level L Leveled Book Word Count: 607Document9 paginiInsect Life Cycle: A Reading A-Z Level L Leveled Book Word Count: 607Bombo MartinÎncă nu există evaluări
- Arecanut: Aesa Based Ipm PackageDocument51 paginiArecanut: Aesa Based Ipm PackageAzizrahman AbubakarÎncă nu există evaluări
- Umi Umd 5867 PDFDocument234 paginiUmi Umd 5867 PDFmicabiologistÎncă nu există evaluări
- Chili Pepper Chapter123Document66 paginiChili Pepper Chapter123Sherry Gesim70% (10)
- AAUA Post Utme Past Questions and Answers PDFDocument121 paginiAAUA Post Utme Past Questions and Answers PDFIshola Joseph OluwakamiyeÎncă nu există evaluări
- The Various Communicative Methods Practiced by Animals in The WildDocument2 paginiThe Various Communicative Methods Practiced by Animals in The WildShopee NguyễnÎncă nu există evaluări
- Chapter - 7 Novel Protein Foods: Alternative Sources of Protein For Human ConsumptionDocument14 paginiChapter - 7 Novel Protein Foods: Alternative Sources of Protein For Human Consumptionscience world100% (1)
- Postharvest Handling Definition of TermsDocument8 paginiPostharvest Handling Definition of TermsApril Ervas100% (1)
- P.4 Science Test Revision QuestionsDocument7 paginiP.4 Science Test Revision QuestionsSean Liam50% (2)
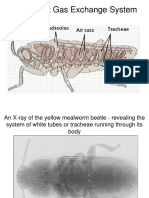
- The Insect Gas Exchange SystemDocument18 paginiThe Insect Gas Exchange SystemkeshavÎncă nu există evaluări
- Viruses As Biological Control AgentsDocument17 paginiViruses As Biological Control AgentsJohn BildanÎncă nu există evaluări
- Wood DestroyingPestStudyGuideDocument58 paginiWood DestroyingPestStudyGuideHeera KardongÎncă nu există evaluări
- Classification of InsecticidesDocument7 paginiClassification of Insecticideshamzashafiq1011Încă nu există evaluări
- Manipulation of Consciousness Sergey Kara MurzaDocument605 paginiManipulation of Consciousness Sergey Kara MurzaOsamaDlikan100% (1)