S-ar putea să vă placă și
- Rejuvenescer Com O Plasma Sanguíneo Dos JovensDe la EverandRejuvenescer Com O Plasma Sanguíneo Dos JovensÎncă nu există evaluări
- Relação Entre Apoptose e Alzheimer - BioquímicaDocument13 paginiRelação Entre Apoptose e Alzheimer - BioquímicagleysonlopesÎncă nu există evaluări
- 4-Cadeia Transportadora de Elétrons e Fosforilação OxidativaDocument49 pagini4-Cadeia Transportadora de Elétrons e Fosforilação OxidativaLudmilla FerreiraÎncă nu există evaluări
- Bioestatística: Guia da Disciplina HíbridaDocument22 paginiBioestatística: Guia da Disciplina HíbridaEmídio Junior DuoÎncă nu există evaluări
- COMPORTAMENTO E BEM-ESTAR DE EQUINOS DE ESPORTEDocument7 paginiCOMPORTAMENTO E BEM-ESTAR DE EQUINOS DE ESPORTEHdms SÎncă nu există evaluări
- Imunologia BásicaDocument6 paginiImunologia BásicaDeborasSouzaÎncă nu există evaluări
- Como a imunidade é adquiridaDocument13 paginiComo a imunidade é adquiridaErick HuebraÎncă nu există evaluări
- Curriculo - Fernando Barbosa CavalcanteDocument4 paginiCurriculo - Fernando Barbosa CavalcanteFernando CavalcanteÎncă nu există evaluări
- Doença, Medicina e Religião na Idade MédiaDocument1 paginăDoença, Medicina e Religião na Idade MédiaSabrina Sales AraujoÎncă nu există evaluări
- Olfato e PaladarDocument16 paginiOlfato e PaladarAmanda NathalyÎncă nu există evaluări
- Introdução À HPLCDocument36 paginiIntrodução À HPLCMarilena MeiraÎncă nu există evaluări
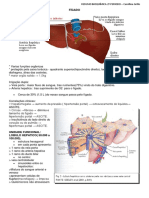
- Bioquímica Do Fígado, Renal e NeurotransmissoresDocument14 paginiBioquímica Do Fígado, Renal e NeurotransmissoresNathália Miranda100% (1)
- Filme retrata descoberta cirúrgica que salvou vidasDocument1 paginăFilme retrata descoberta cirúrgica que salvou vidasLuziane MendonçaÎncă nu există evaluări
- 001 - Introdução A BioquímicaDocument20 pagini001 - Introdução A BioquímicaEdnilton nobre nunesÎncă nu există evaluări
- Bioquímica Fundamentos CelularesDocument9 paginiBioquímica Fundamentos CelularesPriscila CunhaÎncă nu există evaluări
- LeucinoseDocument2 paginiLeucinoseBárbara CostaÎncă nu există evaluări
- Erros inatos do metabolismo: características, manifestações e principais doenças (PKU e galactosemiaDocument20 paginiErros inatos do metabolismo: características, manifestações e principais doenças (PKU e galactosemiaPatrícia AlmeidaÎncă nu există evaluări
- Microbiologia de alimentos: introdução aos microrganismosDocument50 paginiMicrobiologia de alimentos: introdução aos microrganismosPaulo Roberto FilhoÎncă nu există evaluări
- Aula 03 - Genética Quantitativa - Poligenes, Herdabilidade, Ganho de Seleção e Heterose - Sem RespostaDocument67 paginiAula 03 - Genética Quantitativa - Poligenes, Herdabilidade, Ganho de Seleção e Heterose - Sem RespostarakelÎncă nu există evaluări
- Fundamentos Da BioquímicaDocument95 paginiFundamentos Da BioquímicaLéya Mateus100% (1)
- Enzimas do ciclo da ureiaDocument57 paginiEnzimas do ciclo da ureiaPatrícia AlmeidaÎncă nu există evaluări
- Atlas do sedimento urinárioDocument18 paginiAtlas do sedimento urinárioFernanda ZanellaÎncă nu există evaluări
- Livros de MedicinaDocument10 paginiLivros de MedicinasÎncă nu există evaluări
- 1 Processos Biologicos Básicos PDFDocument5 pagini1 Processos Biologicos Básicos PDFrejaneÎncă nu există evaluări
- Parasitoses humanasDocument2 paginiParasitoses humanasOfficecor Corretora DE SegurosÎncă nu există evaluări
- Neurobiologia Dos Comportamentos AditivosDocument39 paginiNeurobiologia Dos Comportamentos AditivosJoão SousaÎncă nu există evaluări
- Genética - O Que Esse Assunto Tem A Ver Com Você?De la EverandGenética - O Que Esse Assunto Tem A Ver Com Você?Încă nu există evaluări
- Relatório: Técnicas de Microscopia E Coloração de Gram: Universidade Estadual de GoiásDocument3 paginiRelatório: Técnicas de Microscopia E Coloração de Gram: Universidade Estadual de Goiásjoyce camposÎncă nu există evaluări
- Biologia e Bioquímica Humana: Dra. Marcia Cristina de Souza Lara KameiDocument184 paginiBiologia e Bioquímica Humana: Dra. Marcia Cristina de Souza Lara KameiYasmin DiasÎncă nu există evaluări
- NUTRILITE Perfil - NutricionalDocument2 paginiNUTRILITE Perfil - NutricionalKaryni da Vila100% (1)
- 01 Introdução À FarmacologiaDocument24 pagini01 Introdução À FarmacologiaBruno Sousa100% (2)
- SíNDROME DE DIGEORGEDocument14 paginiSíNDROME DE DIGEORGEYalli SchettiniÎncă nu există evaluări
- 0-Aula Prática Perfil LipídicoDocument7 pagini0-Aula Prática Perfil LipídicoAllana NegreirosÎncă nu există evaluări
- Química - Carboidratos, Gorduras, Colesterol e ProteínasDocument4 paginiQuímica - Carboidratos, Gorduras, Colesterol e ProteínasQuímica_Moderna100% (3)
- Aula 3. Sistema Nervoso PDFDocument8 paginiAula 3. Sistema Nervoso PDFLiziane RodriguesÎncă nu există evaluări
- Doenças Causadas P Microorganismos de Origem Alimentar Prof Juliana 1Document19 paginiDoenças Causadas P Microorganismos de Origem Alimentar Prof Juliana 1karinaaparecidasanchesbatista100% (1)
- Portfólio DOENÇAS INFECIOSASDocument74 paginiPortfólio DOENÇAS INFECIOSASSandra Lameu100% (1)
- Tecido Conjuntivo EspecializadoDocument11 paginiTecido Conjuntivo EspecializadoGivaldo DamascenoÎncă nu există evaluări
- LiteraturaMedica DIM Di Indol MetanoDocument1 paginăLiteraturaMedica DIM Di Indol MetanoDeonice Sazonov50% (2)
- Reflexos oculares e patelarDocument2 paginiReflexos oculares e patelarFlávia CorcinoÎncă nu există evaluări
- Morfologia e Citologia BacterianaDocument60 paginiMorfologia e Citologia BacterianaFarmacêutica LiteráriaÎncă nu există evaluări
- Roteiro EstagioDocument4 paginiRoteiro EstagioGilson MarinhoÎncă nu există evaluări
- Lista de Exercício Prof Gui PDFDocument6 paginiLista de Exercício Prof Gui PDFMarcela GomesÎncă nu există evaluări
- Boas Práticas em Laboratorio - Biologia PDFDocument270 paginiBoas Práticas em Laboratorio - Biologia PDFAlexandre CasarolliÎncă nu există evaluări
- Introdução à Fisiologia HumanaDocument26 paginiIntrodução à Fisiologia HumanaMarcos MeloÎncă nu există evaluări
- Desenvolvimento embrionárioDocument43 paginiDesenvolvimento embrionárioTalita FreitasÎncă nu există evaluări
- Centro Terapêutico para Tratamento de Depressão e AnsiedadeDocument64 paginiCentro Terapêutico para Tratamento de Depressão e AnsiedadeAlessandra BatalhaÎncă nu există evaluări
- Fisiologia do adulto e envelhecimentoDocument34 paginiFisiologia do adulto e envelhecimentoMaria Regina CamargoÎncă nu există evaluări
- Habilitações BiomedicinaDocument7 paginiHabilitações BiomedicinaÉrika BrandãoÎncă nu există evaluări
- Medicamentos Fracionados - Guia para FarmaceuticosDocument28 paginiMedicamentos Fracionados - Guia para FarmaceuticosdruguniaoÎncă nu există evaluări
- Trabalho de Meiose e MitoseDocument10 paginiTrabalho de Meiose e MitoseNeide Diassis Salomão DavyÎncă nu există evaluări
- Autoanticorpos após uso de terapia biológica: estudo multicêntrico brasileiroDe la EverandAutoanticorpos após uso de terapia biológica: estudo multicêntrico brasileiroÎncă nu există evaluări
- Fabricação do queijo mussarela em 40 passosDocument4 paginiFabricação do queijo mussarela em 40 passosFelipeÎncă nu există evaluări
- 35 Doenças NeurodegenerativasDocument15 pagini35 Doenças NeurodegenerativasFábio Marcelino Duarte DuarteÎncă nu există evaluări
- Estruturas e características da célulaDocument20 paginiEstruturas e características da célulaGILMARA PRADO DE SOUSA100% (2)
- Relatório de Química GeralDocument7 paginiRelatório de Química GeralAldoMaiaÎncă nu există evaluări
- Educação Básica e Suas Práticas de EnsinoDocument76 paginiEducação Básica e Suas Práticas de Ensinomarcoaureliovieira100% (1)
- Edgar Allan Poe - Contos de Terror, Misterio e Morte 05 - MetzengersteinDocument6 paginiEdgar Allan Poe - Contos de Terror, Misterio e Morte 05 - MetzengersteinMensageiro ObscuroÎncă nu există evaluări
- A Morte Que Fez Um Homem RicoDocument4 paginiA Morte Que Fez Um Homem RicoCentro Infantil Justino TeixeiraÎncă nu există evaluări
- Prova de Tanatologia sobre Morte e LutoDocument5 paginiProva de Tanatologia sobre Morte e LutoCamilly PeclyÎncă nu există evaluări
- Escritura de Inventário e Adjudicação Dos Bens Deixados Pelo Falecimento deDocument5 paginiEscritura de Inventário e Adjudicação Dos Bens Deixados Pelo Falecimento deWarllingson Derick100% (1)
- A origem, natureza e destino do homem na morteDocument60 paginiA origem, natureza e destino do homem na morteNadjane DamascenaÎncă nu există evaluări
- Pai Nosso Pedras Vivas cifraDocument1 paginăPai Nosso Pedras Vivas cifraRafael Rangel FagundesÎncă nu există evaluări
- Afogamento em 40Document23 paginiAfogamento em 40AanÎncă nu există evaluări
- Elder Ferreira Da Fonseca Até João BeleriqueDocument2 paginiElder Ferreira Da Fonseca Até João BeleriqueElderFerreiraÎncă nu există evaluări
- As Marcas Do Autor em o Ano Da Morte de Ricardo ReisDocument42 paginiAs Marcas Do Autor em o Ano Da Morte de Ricardo ReisMaria De Jesus BezerraÎncă nu există evaluări
- Aula 01 - Crimes Contra PessoaDocument10 paginiAula 01 - Crimes Contra PessoaVictor ParasinÎncă nu există evaluări
- Cânticos de umbandaDocument2 paginiCânticos de umbandaJoao Noronha EmilianoÎncă nu există evaluări
- Acidentes Na InfânciaDocument91 paginiAcidentes Na InfânciajuliaperezÎncă nu există evaluări
- Cantiga da barataDocument13 paginiCantiga da barataAnaAnjosÎncă nu există evaluări
- TCC Controle de Pragas UrbanasDocument33 paginiTCC Controle de Pragas UrbanasPatricia Cedraz100% (2)
- Naomi Lucas - 02 - Minotaur Prayer (Rev)Document287 paginiNaomi Lucas - 02 - Minotaur Prayer (Rev)daniely pacheco100% (2)
- Entomologia Forense: estudo de insetos na determinação da data da morteDocument32 paginiEntomologia Forense: estudo de insetos na determinação da data da morteTelma PzybylskiÎncă nu există evaluări
- Medicina Legal TANATOLOGIA ForenseDocument13 paginiMedicina Legal TANATOLOGIA ForenseElen Vasconcellos63% (8)
- Aula Entomologia Forense ThyssDocument14 paginiAula Entomologia Forense ThyssDiego AssisÎncă nu există evaluări
- Direito Penal III - Crimes Contra a PessoaDocument59 paginiDireito Penal III - Crimes Contra a PessoaSelma OliveiraÎncă nu există evaluări
- A Bailarina Da MorteDocument2 paginiA Bailarina Da MorteAnonymous TQbS7BVZCÎncă nu există evaluări
- Novo Alfabetização Com Boquinhas Vol 5Document57 paginiNovo Alfabetização Com Boquinhas Vol 5Piti Kersting100% (1)
- Introduçã1Document2 paginiIntroduçã1RICARDOÎncă nu există evaluări
- Processador de texto - Funcionalidades avançadasDocument5 paginiProcessador de texto - Funcionalidades avançadasrflp87Încă nu există evaluări
- Epidemiologia Nutrição UFRGSDocument22 paginiEpidemiologia Nutrição UFRGSAmanda Gomes100% (1)
- Soprano SoloDocument2 paginiSoprano SoloCleyton CruzÎncă nu există evaluări
- Apostila 315 MensagensDocument315 paginiApostila 315 MensagensjessylaneeÎncă nu există evaluări
- Comentário: Domingo de RAMOS - Ano ADocument2 paginiComentário: Domingo de RAMOS - Ano AJosé Ribeiro de LimaÎncă nu există evaluări
- FichaAvaliacaoo Ano Da Morte de RRDocument7 paginiFichaAvaliacaoo Ano Da Morte de RRMARIAJBEZERRAÎncă nu există evaluări
- O Poderoso Orixá OgunDocument4 paginiO Poderoso Orixá Ogunmacelovieira2009Încă nu există evaluări
- Certidao Obito Mae PDFDocument1 paginăCertidao Obito Mae PDFKerlon D. SilveiraÎncă nu există evaluări