S-ar putea să vă placă și
- Genetics, Lecture 11 (Lecture Notes)Document16 paginiGenetics, Lecture 11 (Lecture Notes)Ali Al-QudsiÎncă nu există evaluări
- This Video: To Do: Watch To Complete This TableDocument1 paginăThis Video: To Do: Watch To Complete This TableLaura Cormane100% (1)
- Developmental Biology: Understanding Normal and Abnormal DevelopmentDocument17 paginiDevelopmental Biology: Understanding Normal and Abnormal DevelopmentChetan GopalÎncă nu există evaluări
- The Structure and Function of Animal Cell Components: An Introductory TextDe la EverandThe Structure and Function of Animal Cell Components: An Introductory TextÎncă nu există evaluări
- Developmental BiologyDocument21 paginiDevelopmental BiologygahanÎncă nu există evaluări
- 3 Genetics: 3.1 GenesDocument22 pagini3 Genetics: 3.1 Genesapi-285490927100% (1)
- Consolidated School Quality ReportDocument108 paginiConsolidated School Quality ReportSudhir VashistÎncă nu există evaluări
- Innate ImmunityDocument9 paginiInnate Immunitynascha dumpÎncă nu există evaluări
- Structures of A Virus: A Tour To The Nature's Deadliest WeaponDocument72 paginiStructures of A Virus: A Tour To The Nature's Deadliest WeaponcezalynÎncă nu există evaluări
- Cellular Respiration 1Document28 paginiCellular Respiration 1Maria Flor PabeloniaÎncă nu există evaluări
- Intermediate FilamentsDocument4 paginiIntermediate FilamentsSai SridharÎncă nu există evaluări
- Lecture Outline: Chapter 5 The Structure and Function of MacromoleculesDocument12 paginiLecture Outline: Chapter 5 The Structure and Function of MacromoleculesSanvir RulezzÎncă nu există evaluări
- Mechanism of EvolutionDocument41 paginiMechanism of EvolutionChristian EstradaÎncă nu există evaluări
- DNA Structure and ReplicationDocument12 paginiDNA Structure and ReplicationJhune Dominique GalangÎncă nu există evaluări
- 8.lysosomes, Structure and FunctionDocument50 pagini8.lysosomes, Structure and Functionkubuldinho100% (3)
- Cell Biology LaboratoryDocument319 paginiCell Biology LaboratoryNazaqat FarooqÎncă nu există evaluări
- Welcome All !... : Presented By: Dr. Nidhi SrivastavaDocument55 paginiWelcome All !... : Presented By: Dr. Nidhi SrivastavaNivedita DasÎncă nu există evaluări
- Molecular GeneticsDocument3 paginiMolecular GeneticsPRINTDESK by DanÎncă nu există evaluări
- Information Brochure: Department of Genetics, Immunology and Biochemistry Maharashtra University of Health SciencesDocument16 paginiInformation Brochure: Department of Genetics, Immunology and Biochemistry Maharashtra University of Health SciencesSushant JadhavÎncă nu există evaluări
- Pedigree AnalysisDocument37 paginiPedigree Analysisshelren onlagadaÎncă nu există evaluări
- BIOL-1110 - Unit 3 - Study GuideDocument3 paginiBIOL-1110 - Unit 3 - Study GuideOmarÎncă nu există evaluări
- Microbiology+simple+notes .Nursing EduTechDocument111 paginiMicrobiology+simple+notes .Nursing EduTechjmbrand jmÎncă nu există evaluări
- IntroductionDocument24 paginiIntroductioninoka911Încă nu există evaluări
- Genetics SyllabusDocument7 paginiGenetics SyllabusAnonymous q6VpovV6pTÎncă nu există evaluări
- Plant Taxonomy NotesDocument38 paginiPlant Taxonomy Notesopolla nianorÎncă nu există evaluări
- Genetics LectureDocument30 paginiGenetics Lectureapi-295463484Încă nu există evaluări
- Organization and Expression of Immunoglobulin GenesDocument3 paginiOrganization and Expression of Immunoglobulin GenesDavid CastilloÎncă nu există evaluări
- Receptors in the Evolution and Development of the Brain: Matter into MindDe la EverandReceptors in the Evolution and Development of the Brain: Matter into MindÎncă nu există evaluări
- The Pyridine Nucleotide CoenzymesDe la EverandThe Pyridine Nucleotide CoenzymesJohannes EverseÎncă nu există evaluări
- The Plasma Proteins V5: Structure, Function, and Genetic ControlDe la EverandThe Plasma Proteins V5: Structure, Function, and Genetic ControlFrank PutnamEvaluare: 3 din 5 stele3/5 (1)
- Biology Lecture, Chapter 5Document84 paginiBiology Lecture, Chapter 5Nick GoldingÎncă nu există evaluări
- Phylogenetic Tree: Tree Is A Branching Diagram or "Tree"Document6 paginiPhylogenetic Tree: Tree Is A Branching Diagram or "Tree"Juan Santiago Pineda Rodriguez PequeÎncă nu există evaluări
- Cell Biology A Comprehensive Treatise V4: Gene Expression: Translation and the Behavior of ProteinsDe la EverandCell Biology A Comprehensive Treatise V4: Gene Expression: Translation and the Behavior of ProteinsÎncă nu există evaluări
- Molecular Modelling and Drug DesigningDocument16 paginiMolecular Modelling and Drug DesigningAsh-shirHasanÎncă nu există evaluări
- Cellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsDe la EverandCellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsCharles G. CochraneÎncă nu există evaluări
- Bio 244 Chapter 10 NotesDocument7 paginiBio 244 Chapter 10 NotesEscabarte Abkilan Cyrian GrioÎncă nu există evaluări
- DNA Replication Sequencing Revision 2017Document58 paginiDNA Replication Sequencing Revision 2017Daniel sifuentes garciaÎncă nu există evaluări
- CellDocument47 paginiCellAnami NizamÎncă nu există evaluări
- Monoclonal Antibodies Against Bacteria: Volume IIIDe la EverandMonoclonal Antibodies Against Bacteria: Volume IIIAlberto J. L. MacarioÎncă nu există evaluări
- Dna and Rna Powerpoint 2Document46 paginiDna and Rna Powerpoint 2api-267309851Încă nu există evaluări
- Cell and Molecular BiologyDocument4 paginiCell and Molecular BiologyRea Joyce AldefollaÎncă nu există evaluări
- PAM Abd BLOSUMDocument3 paginiPAM Abd BLOSUMhohoiyinÎncă nu există evaluări
- Genetic Engineering (Gowshia, Fatimah, Inthuja)Document15 paginiGenetic Engineering (Gowshia, Fatimah, Inthuja)Lacramioara Amalia Amariei100% (1)
- Genetics and Cytogenetics For UndergradsDocument145 paginiGenetics and Cytogenetics For UndergradsHarsh vardhanÎncă nu există evaluări
- OpenGeneticsLectures Fall2017Document405 paginiOpenGeneticsLectures Fall2017JustinVoÎncă nu există evaluări
- Behavioural Science Notes UpadetedDocument142 paginiBehavioural Science Notes UpadetedKALAMYA BRIAN BWAYOÎncă nu există evaluări
- Cell Biology Practice 2Document15 paginiCell Biology Practice 2NgMinhHaiÎncă nu există evaluări
- Linkage: Harshraj Subhash Shinde KKW, Cabt, NashikDocument14 paginiLinkage: Harshraj Subhash Shinde KKW, Cabt, Nashiksivaram888Încă nu există evaluări
- Reproduction and Sexuality in Marine Fishes: Patterns and ProcessesDe la EverandReproduction and Sexuality in Marine Fishes: Patterns and ProcessesKathleen S. ColeÎncă nu există evaluări
- To Genetics: Eighth EditionDocument41 paginiTo Genetics: Eighth EditionNikkoSilvaÎncă nu există evaluări
- CH 13 Lecture PresentationDocument97 paginiCH 13 Lecture PresentationDalia M. MohsenÎncă nu există evaluări
- Antigen Processing Process and MHC AssociationDocument3 paginiAntigen Processing Process and MHC AssociationMahathir Mohmed100% (2)
- Mechanisms of Hormone Action NotesDocument3 paginiMechanisms of Hormone Action Notesapi-390361165Încă nu există evaluări
- BIOL223-Lab 7Document40 paginiBIOL223-Lab 7chicken fries100% (2)
- VirusDocument57 paginiVirusSophan HadieÎncă nu există evaluări
- Cloning Dolly & MicromanipulationDocument29 paginiCloning Dolly & Micromanipulationnitinyadav16Încă nu există evaluări
- Crispr Cas HajarDocument21 paginiCrispr Cas HajarHajira Fatima100% (1)
- Spermatogenesis: An Overview: Rakesh Sharma and Ashok AgarwalDocument25 paginiSpermatogenesis: An Overview: Rakesh Sharma and Ashok Agarwalumar umarÎncă nu există evaluări
- BIO50-Course Specification-AY2022-2023-Term2-V1.0 PDFDocument6 paginiBIO50-Course Specification-AY2022-2023-Term2-V1.0 PDFD M HÎncă nu există evaluări
- Sea Urchin EmbryologyDocument6 paginiSea Urchin EmbryologyAmy HollingsworthÎncă nu există evaluări
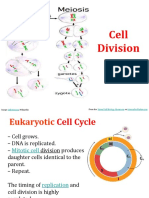
- Review of Cell Division BSC StudentsDocument64 paginiReview of Cell Division BSC StudentsKripa SusanÎncă nu există evaluări
- Science Module 2 1Document18 paginiScience Module 2 1Mark Alexis P. RimorinÎncă nu există evaluări
- CH 11 PPT Cell Communication 1Document77 paginiCH 11 PPT Cell Communication 1api-270681964Încă nu există evaluări
- Cell DivisionDocument17 paginiCell DivisionRaj AgrawalÎncă nu există evaluări
- Q1 Week 5 Biology G9Document17 paginiQ1 Week 5 Biology G9Christopher MarindoqueÎncă nu există evaluări
- Igenetics Pearson New International Edition A Molecular Approach Ebook PDFDocument41 paginiIgenetics Pearson New International Edition A Molecular Approach Ebook PDFkathleen.watts301100% (38)
- Student Exploration: Cell Division: Vocabulary: Cell Division, Centriole, Centromere, Chromatid, Chromatin, ChromosomeDocument4 paginiStudent Exploration: Cell Division: Vocabulary: Cell Division, Centriole, Centromere, Chromatid, Chromatin, ChromosomerosieÎncă nu există evaluări
- 9700 w22 Ms 42 PDFDocument19 pagini9700 w22 Ms 42 PDFManha KhalidÎncă nu există evaluări
- Mitosis-And-Meiosis-Ppt 7781Document30 paginiMitosis-And-Meiosis-Ppt 7781zhao YingingÎncă nu există evaluări
- Characteristics of Living Things Lecture NotesDocument12 paginiCharacteristics of Living Things Lecture NotesErica HartÎncă nu există evaluări
- CH 12 Reading GuideDocument9 paginiCH 12 Reading GuideKapil NathanÎncă nu există evaluări
- Biology Assignment (Stem Cells)Document3 paginiBiology Assignment (Stem Cells)Dwayne CoelhoÎncă nu există evaluări
- Ncert Solutions Jan27 For Class 7 Science Chapter 12 Reproduction in PlantsDocument5 paginiNcert Solutions Jan27 For Class 7 Science Chapter 12 Reproduction in PlantsPRATIK SONWANEÎncă nu există evaluări
- Jyoti Singh and Anuj KumarDocument11 paginiJyoti Singh and Anuj Kumarudayakundu143Încă nu există evaluări
- Protein Synthesis Simulation ActivityDocument4 paginiProtein Synthesis Simulation ActivitySHARIFAH BINTI HASSAN MoeÎncă nu există evaluări
- S10 Q3 Enhanced Hybrid Module 4 Week 4 FinalDocument16 paginiS10 Q3 Enhanced Hybrid Module 4 Week 4 FinalBriannah loiuse P. AdalidÎncă nu există evaluări
- Matdid 207367Document40 paginiMatdid 207367Deblina JanaÎncă nu există evaluări
- Cell PhysiologyDocument7 paginiCell PhysiologyReslie Joy Serrano100% (1)
- Genetika - Tautan Dan Pemetaan KromosomDocument21 paginiGenetika - Tautan Dan Pemetaan KromosomKhairil MikdarÎncă nu există evaluări
- Cancer Cell Development - Lecture Day 5Document17 paginiCancer Cell Development - Lecture Day 5blakeÎncă nu există evaluări
- BIOL223-Lab 7Document40 paginiBIOL223-Lab 7chicken fries100% (2)
- PDFDocument4 paginiPDFYordan Gomez Capellan0% (1)
- STPM Term 3 BiologyDocument9 paginiSTPM Term 3 BiologyYeap Chee BengÎncă nu există evaluări
- Stage 8 - Chapter 3 WorksheetsDocument3 paginiStage 8 - Chapter 3 WorksheetsMoath AlsaidÎncă nu există evaluări
- Revision Ch-Ii (Sexual Reproduction in Flowering Plants)Document11 paginiRevision Ch-Ii (Sexual Reproduction in Flowering Plants)Sahil GautamÎncă nu există evaluări
- 6 Answers To End-Of-Chapter QuestionsDocument6 pagini6 Answers To End-Of-Chapter Questionsalyss ramdathÎncă nu există evaluări
- Reproduction Extra QuestionsDocument10 paginiReproduction Extra Questionschayyakudopa9Încă nu există evaluări