S-ar putea să vă placă și
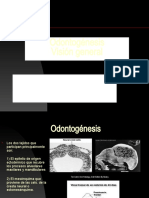
- ODONTOGENESISDocument57 paginiODONTOGENESISEstephania PradoÎncă nu există evaluări
- Odontogenesis RevolutionDocument59 paginiOdontogenesis Revolutionpaolahermosa100% (4)
- Odontogénesis: Formación de los DientesDocument13 paginiOdontogénesis: Formación de los DientesKaren Ortiz100% (1)
- OdontogénesisDocument7 paginiOdontogénesisThomas PazÎncă nu există evaluări
- Embriologia Dentaria OdontogenesisDocument12 paginiEmbriologia Dentaria OdontogenesisDarío Neira100% (1)
- Morfogénesis Del Órgano DentarioDocument4 paginiMorfogénesis Del Órgano DentarioAripcy MatiasÎncă nu există evaluări
- Odontogenesis Resumen Histologia y Embriologia BucodentalDocument13 paginiOdontogenesis Resumen Histologia y Embriologia BucodentalAlejandra NietoÎncă nu există evaluări
- Articulo OdontogenesisDocument20 paginiArticulo OdontogenesisMinerva de SparrowÎncă nu există evaluări
- Complejo Dentino PulparDocument6 paginiComplejo Dentino PulparAremi SánchezÎncă nu există evaluări
- Odontogenesis ..1Document21 paginiOdontogenesis ..1Hernán Murillo Chablé0% (1)
- Odonto GenesisDocument38 paginiOdonto GenesisNaturall KileerÎncă nu există evaluări
- OdontogénesisDocument18 paginiOdontogénesistriplepe100% (1)
- MucosaBucal40Document82 paginiMucosaBucal40aaronÎncă nu există evaluări
- Ciclo Vital Del OdontoblastoDocument6 paginiCiclo Vital Del OdontoblastoIsaac Medina0% (1)
- Histologia Del HuesoDocument6 paginiHistologia Del HuesoAmaiaTelletxeaGortazarÎncă nu există evaluări
- Ligamento PeriodontalDocument14 paginiLigamento PeriodontalIsabella VC100% (1)
- OdontogénesisDocument17 paginiOdontogénesisdmolinagÎncă nu există evaluări
- PeriodontoDocument21 paginiPeriodontoKariina Cuxiim'Încă nu există evaluări
- OdontogénesisDocument39 paginiOdontogénesischewa500100% (2)
- El Esmalte DentalDocument22 paginiEl Esmalte DentalAndrea Ramon nuñezÎncă nu există evaluări
- Inervación de Cabeza y DientesDocument32 paginiInervación de Cabeza y DientesDEYSI EDITH GUERRERO NEYRAÎncă nu există evaluări
- La EncíaDocument47 paginiLa Encíamelisa3mbÎncă nu există evaluări
- Desarrollo embrionario de los dientes y periodontoDocument7 paginiDesarrollo embrionario de los dientes y periodontoYessica JimenezÎncă nu există evaluări
- Anatomia Glandulas SalivalesDocument17 paginiAnatomia Glandulas SalivalesLourdes PalaciosÎncă nu există evaluări
- AMELOGÉNESISDocument21 paginiAMELOGÉNESISJhonatan Chamorro CervantesÎncă nu există evaluări
- Embriologia Dentaria OdontogenesisDocument34 paginiEmbriologia Dentaria OdontogenesisLaura LeoÎncă nu există evaluări
- PULPA DENTARIA Banco de ApuntesDocument8 paginiPULPA DENTARIA Banco de ApuntesKarinaÎncă nu există evaluări
- Tipos de DentinaDocument4 paginiTipos de DentinafernandoÎncă nu există evaluări
- Embriohistología de la cavidad bucalDocument12 paginiEmbriohistología de la cavidad bucalKarina ReyesÎncă nu există evaluări
- Hueso AlveolarDocument18 paginiHueso AlveolarGerardo Martínez75% (4)
- Operatoria DentalDocument12 paginiOperatoria DentalKathiinha MeeaawÎncă nu există evaluări
- Mucosa OralDocument3 paginiMucosa OralRichard CarÎncă nu există evaluări
- Quiste NasopalatinoDocument8 paginiQuiste NasopalatinoEliana MejiaÎncă nu există evaluări
- Caries DentalDocument5 paginiCaries Dentalajhm89Încă nu există evaluări
- Nervio Alveolar InferiorDocument11 paginiNervio Alveolar InferiorLisie Margott Quispe PonceÎncă nu există evaluări
- Complejo Dentino PulparDocument61 paginiComplejo Dentino PulparCrissia Palomino BediaÎncă nu există evaluări
- Hueso Alveolar IDocument8 paginiHueso Alveolar IFiorellaOlivaresAloneinthislifeÎncă nu există evaluări
- Embriogenesis PeriodontoDocument19 paginiEmbriogenesis PeriodontoSarita GaleanoÎncă nu există evaluări
- Complejo Dentino PulparDocument20 paginiComplejo Dentino PulparDennis MejiaÎncă nu există evaluări
- Embriogenesis Dental FranchescaDocument27 paginiEmbriogenesis Dental FranchescaHugoRicardo100% (1)
- Encia PDFDocument17 paginiEncia PDFCesar Omar VasquezÎncă nu există evaluări
- Formacion Del Hueso en NinosDocument5 paginiFormacion Del Hueso en Ninosanon_571797493Încă nu există evaluări
- Anatomía y características de la encíaDocument71 paginiAnatomía y características de la encíaCarlos Ugalde Iglesias100% (1)
- Ligamento Periodontal MMMMMDocument5 paginiLigamento Periodontal MMMMMana100% (1)
- Fibroma Cementificante - OsificanteDocument8 paginiFibroma Cementificante - OsificantePucho Gregorio Caballero PonceÎncă nu există evaluări
- Condiciones No Patológicas de La Cavidad BucalDocument6 paginiCondiciones No Patológicas de La Cavidad BucalJoseLuisEspinozaBeltranÎncă nu există evaluări
- Fisiología PulparDocument7 paginiFisiología Pulparlalospa4Încă nu există evaluări
- Lesiones PulparesDocument107 paginiLesiones PulparesfabrizioaryanÎncă nu există evaluări
- Cementos DentalesDocument7 paginiCementos DentalesPauli Morales100% (2)
- 6 Fluido GingivalDocument17 pagini6 Fluido GingivalDana Lisett Ygnacio LlanosÎncă nu există evaluări
- Hueso Alveolar, Ligamento y CementoDocument28 paginiHueso Alveolar, Ligamento y Cementoambar100% (1)
- PeriodontoDocument35 paginiPeriodontoMiguel Soni CasillasÎncă nu există evaluări
- Histología de la erupción dentalDocument33 paginiHistología de la erupción dentalHISTOLOGIA FOUCÎncă nu există evaluări
- Cemento y HuesoDocument68 paginiCemento y HuesoJhónatan Vilca ChaparroÎncă nu există evaluări
- Formación del Germen DentarioDocument4 paginiFormación del Germen DentarioMario Morales RiveraÎncă nu există evaluări
- Atm Histologia Bucal-20170404-12172799Document65 paginiAtm Histologia Bucal-20170404-12172799Estefania RodriguezÎncă nu există evaluări
- Cuidados en salud bucal para personas con discapacidad sensorialDe la EverandCuidados en salud bucal para personas con discapacidad sensorialÎncă nu există evaluări
- Prácticas de anatomía con enfoque odontológicoDe la EverandPrácticas de anatomía con enfoque odontológicoÎncă nu există evaluări
- Desarrollo dental: Odontogénesis y morfogénesisDocument20 paginiDesarrollo dental: Odontogénesis y morfogénesisBosco ReyesÎncă nu există evaluări
- Vih SidaDocument32 paginiVih Sidamelrivmar100% (5)
- Coccidioides ImmitisDocument11 paginiCoccidioides ImmitisCarlos Baas AkeÎncă nu există evaluări
- Dril y Encaje - Diana BlayneDocument282 paginiDril y Encaje - Diana Blaynetallyslucia0% (1)
- 22-Anexo B. Las CerditasDocument22 pagini22-Anexo B. Las CerditasElsa Alcala0% (1)
- Haemonchus ContortusDocument19 paginiHaemonchus ContortusKyoÎncă nu există evaluări
- Anatomia Del CaracolDocument3 paginiAnatomia Del CaracolMIGUEZ ANDRADE FABIOLA CAROLINAÎncă nu există evaluări
- Cuaderno Caza LobatosDocument177 paginiCuaderno Caza LobatosJohn CooperÎncă nu există evaluări
- Robinson Crusoe El Viaje Del Beagle - ReglasDocument23 paginiRobinson Crusoe El Viaje Del Beagle - Reglasmpezoa.aÎncă nu există evaluări
- Ejemplos de Sustantivos ComunesDocument2 paginiEjemplos de Sustantivos ComunesJuan Pablo JolomnáÎncă nu există evaluări
- Eleanor Woods - Mi Único Amante (Destierro Amoroso)Document85 paginiEleanor Woods - Mi Único Amante (Destierro Amoroso)Solange Cifuentes80% (5)
- Monstruos y Seres Fantásticos de La Mitología GriegaDocument3 paginiMonstruos y Seres Fantásticos de La Mitología GriegaMariela ZepolÎncă nu există evaluări
- Infecciones GinecologicasDocument61 paginiInfecciones GinecologicascarlagarciagyoÎncă nu există evaluări
- Ejercicios. PsicofisiologiaDocument6 paginiEjercicios. PsicofisiologiaYensi Fernanda Delgado CaicedoÎncă nu există evaluări
- CAPILARIASISDocument13 paginiCAPILARIASISYune DbÎncă nu există evaluări
- Los EuriptéridosDocument2 paginiLos EuriptéridosLuis Fidel Raimi RevillaÎncă nu există evaluări
- Odontología Restauradora en Dientes DeciduosDocument9 paginiOdontología Restauradora en Dientes DeciduosJuan Carlos PinedaÎncă nu există evaluări
- Los Orishas Sus Animales y OfrendasDocument11 paginiLos Orishas Sus Animales y OfrendasAnonymous 8PgitGÎncă nu există evaluări
- Farid DieckDocument3 paginiFarid Dieckximegonza034Încă nu există evaluări
- Patricio WebquestDocument16 paginiPatricio WebquestKarinaÎncă nu există evaluări
- Problemas de Decimales-5°-20Document12 paginiProblemas de Decimales-5°-20EDGAR HIGUERA RAMOSÎncă nu există evaluări
- Caderno de Caligrafia e SilabasDocument30 paginiCaderno de Caligrafia e SilabasHomeschooling RegistrosÎncă nu există evaluări
- SILVA, TAREA N°1 Eutanasia A Los Perros Asilvestrados en Áreas de La BiodiversidadDocument4 paginiSILVA, TAREA N°1 Eutanasia A Los Perros Asilvestrados en Áreas de La BiodiversidadFelipe Silva RiquelmeÎncă nu există evaluări
- El Mito de La Creacion Del UniversoDocument1 paginăEl Mito de La Creacion Del UniversoDaniel PovedaÎncă nu există evaluări
- Cesar Vallejo - Heces (Poema Analizado)Document3 paginiCesar Vallejo - Heces (Poema Analizado)David Fernandez100% (1)
- Piel HistologiaDocument8 paginiPiel HistologiaSamm Sango ArteagaÎncă nu există evaluări
- Ficha Las Celulas para Quinto de PrimariaDocument2 paginiFicha Las Celulas para Quinto de PrimariaNorma FuentesÎncă nu există evaluări
- Tarea 4 Comu2Document3 paginiTarea 4 Comu2Yasmin Malpartida Rodriguez75% (12)
- Trabajo Grupal 2Document12 paginiTrabajo Grupal 2Mafer CastroÎncă nu există evaluări
- Area de Hematología: Empresa: Salud Total EPS-S S.ADocument2 paginiArea de Hematología: Empresa: Salud Total EPS-S S.ACONTADOR CONTADORÎncă nu există evaluări
- Normas ApaDocument13 paginiNormas ApaNikolhija Sinisterra0% (1)