S-ar putea să vă placă și
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (119)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Full Specification and Technical Data Sheet of Potato Dextrose AgarDocument2 paginiFull Specification and Technical Data Sheet of Potato Dextrose AgarKunal VermaÎncă nu există evaluări
- International Rice Research Notes Vol.28 No.1Document80 paginiInternational Rice Research Notes Vol.28 No.1ccquintosÎncă nu există evaluări
- Primescript™ Reverse Transcriptase: Code No. 2680Q Size: 2,000 Units Shipping at 20 Store at 20Document2 paginiPrimescript™ Reverse Transcriptase: Code No. 2680Q Size: 2,000 Units Shipping at 20 Store at 20Isaac Nicholas NotorioÎncă nu există evaluări
- Farkom Jenis ObatDocument3 paginiFarkom Jenis ObatVera KemaraÎncă nu există evaluări
- 1994 AP BioDocument26 pagini1994 AP Bioppphre100% (2)
- IB Biology Notes - 1 Working With DataDocument1 paginăIB Biology Notes - 1 Working With DataWanda Wawa EvirhaÎncă nu există evaluări
- ToxocaraDocument313 paginiToxocaradiana2_0_0_2Încă nu există evaluări
- WizardR Genomic DNA Purification Kit Quick Protocol PDFDocument4 paginiWizardR Genomic DNA Purification Kit Quick Protocol PDFYuliandini Pangestika WiyonoÎncă nu există evaluări
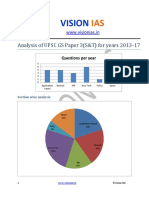
- Analysis of Upsc Gs Paper 3 Science and Tech Without ApproachDocument2 paginiAnalysis of Upsc Gs Paper 3 Science and Tech Without ApproachJames KnotÎncă nu există evaluări
- Stomatal Variation in Dicot and Monocots PDFDocument5 paginiStomatal Variation in Dicot and Monocots PDFReinaur Signey AluningÎncă nu există evaluări
- Biosimilares 2.0Document8 paginiBiosimilares 2.0Carlos Julio Nova LopezÎncă nu există evaluări
- Linkage and Crossing Over ExplainedDocument8 paginiLinkage and Crossing Over ExplainedMahendra singh RajpurohitÎncă nu există evaluări
- Form Pencatatan Vaksin Bri 9 Okt 2021 Di Mcu RSSVDocument211 paginiForm Pencatatan Vaksin Bri 9 Okt 2021 Di Mcu RSSVDeby NoÎncă nu există evaluări
- Bigdye Terminator Protocol v1.1Document74 paginiBigdye Terminator Protocol v1.1Citrawati Dyah Kencono WunguÎncă nu există evaluări
- Antimicrobial Resistance Patterns Among Acinetobacter Baumannii Isolated From Burn Intensive Care Unit in Tripoli, LibyDocument5 paginiAntimicrobial Resistance Patterns Among Acinetobacter Baumannii Isolated From Burn Intensive Care Unit in Tripoli, LibyInternational Medical PublisherÎncă nu există evaluări
- A Plasmid Is A Small DNA Molecule Within A Cell That Is Physically Separated From A Chromosomal DNA and Can Replicate IndependentlyDocument5 paginiA Plasmid Is A Small DNA Molecule Within A Cell That Is Physically Separated From A Chromosomal DNA and Can Replicate Independentlyyaqoob008Încă nu există evaluări
- Lab Act 1Document8 paginiLab Act 1KarenÎncă nu există evaluări
- Dhiraj Kumar, Chengliang Gong-Trends in Insect Molecular Biology and Biotechnology-Springer International Publishing (2018) PDFDocument376 paginiDhiraj Kumar, Chengliang Gong-Trends in Insect Molecular Biology and Biotechnology-Springer International Publishing (2018) PDFAndres Felipe Arias MosqueraÎncă nu există evaluări
- The Pharmaceutical and Chemical JournalDocument2 paginiThe Pharmaceutical and Chemical JournalAlexandru CroitoruÎncă nu există evaluări
- Introduction To MetagenomicsDocument16 paginiIntroduction To MetagenomicsSuhail PatharvatÎncă nu există evaluări
- Microbiology SyllabusDocument56 paginiMicrobiology SyllabusYaswanth SinhaÎncă nu există evaluări
- Nanobiotechnology and BionanotechnologyDocument32 paginiNanobiotechnology and Bionanotechnologysajjad hussainÎncă nu există evaluări
- Raharris - Illumina Infinium 450K ArrayDocument10 paginiRaharris - Illumina Infinium 450K ArraykeyeohÎncă nu există evaluări
- Mfhpb07 Eng ListeriaDocument12 paginiMfhpb07 Eng ListeriaMarce LopezÎncă nu există evaluări
- EU Pharma Draft2Document1 paginăEU Pharma Draft2মোঃ এমদাদুল হকÎncă nu există evaluări
- Principles of Inheritance N Variation PDFDocument4 paginiPrinciples of Inheritance N Variation PDFshodhan shettyÎncă nu există evaluări
- 5e Plan-Central DogmaDocument2 pagini5e Plan-Central Dogmaapi-307962259Încă nu există evaluări
- Gast Caron 2001 Photosymbiotic Associations Planktic Forams RadiolariaDocument7 paginiGast Caron 2001 Photosymbiotic Associations Planktic Forams RadiolariaforaminsÎncă nu există evaluări
- Biomedical Science BrochureDocument12 paginiBiomedical Science BrochuretanasedanielaÎncă nu există evaluări
- MIST M.Sc. Admission-2016Document3 paginiMIST M.Sc. Admission-2016Sajid67% (3)