S-ar putea să vă placă și
- Researches on the Visual Organs of the TrilobitesDe la EverandResearches on the Visual Organs of the TrilobitesÎncă nu există evaluări
- Epinasty and Thermonasty Are Examples of NasticDocument2 paginiEpinasty and Thermonasty Are Examples of NasticHamdan FatahÎncă nu există evaluări
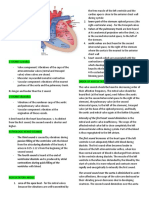
- Anatomy of The HeartDocument3 paginiAnatomy of The HeartCanan YilmazÎncă nu există evaluări
- Will The Real Ventricular Architecture Please StanDocument13 paginiWill The Real Ventricular Architecture Please Stannilberto2Încă nu există evaluări
- Anatomical Variations of The Coronary ArteriesDocument14 paginiAnatomical Variations of The Coronary ArteriesMirza SmajlovićÎncă nu există evaluări
- Changes Lungs Plain Radiograph: Vascular The The of The ChestDocument8 paginiChanges Lungs Plain Radiograph: Vascular The The of The Chestpunct_org3256Încă nu există evaluări
- Sistema de ConduccionDocument5 paginiSistema de Conduccionapi-543828470Încă nu există evaluări
- Visceral OMT: AAO Convocation March 2018 Kenneth Lossing DODocument59 paginiVisceral OMT: AAO Convocation March 2018 Kenneth Lossing DODiana SchlittlerÎncă nu există evaluări
- Health Assessment Semi Finals NotesDocument43 paginiHealth Assessment Semi Finals NotesDazell VarronÎncă nu există evaluări
- Normal and Abnormal AnatomyDocument28 paginiNormal and Abnormal AnatomyTri RachmadijantoÎncă nu există evaluări
- Atrial SeptumDocument7 paginiAtrial SeptumCARLOS ANDRES ALVAREZ TAMARAÎncă nu există evaluări
- Oxygen Saturation Is The: Medicine HemoglobinDocument6 paginiOxygen Saturation Is The: Medicine Hemoglobinsukses belajarÎncă nu există evaluări
- Lecture Notes CVSDocument8 paginiLecture Notes CVSlubnaÎncă nu există evaluări
- Ann Bot-1965-EVANS-205-17Document13 paginiAnn Bot-1965-EVANS-205-17Vrushali PatilÎncă nu există evaluări
- Mizer Es 1963Document11 paginiMizer Es 1963Luca ClementinoÎncă nu există evaluări
- Cardiovascular ProblemsDocument15 paginiCardiovascular ProblemsMYLENE GRACE ELARCOSAÎncă nu există evaluări
- PAPER3 Bahan ReviewDocument5 paginiPAPER3 Bahan ReviewAnis Mebel SikkojangÎncă nu există evaluări
- Pressure-Volume Relationships in Right Ventricle: by Robert R. Lafontant, B.S., Harold Feinberg, PH.D.Document3 paginiPressure-Volume Relationships in Right Ventricle: by Robert R. Lafontant, B.S., Harold Feinberg, PH.D.Kate udeshiÎncă nu există evaluări
- Smith1988 Ventricular InterdependenceDocument15 paginiSmith1988 Ventricular InterdependenceaddaimuhamadfajriÎncă nu există evaluări
- DiaghragmDocument8 paginiDiaghragmSara RaihanÎncă nu există evaluări
- Diaphragm Gross AnatomyDocument3 paginiDiaphragm Gross AnatomyCoy NuñezÎncă nu există evaluări
- Landmarks in The Anatomical Study of The Blood Supply of The SkinDocument11 paginiLandmarks in The Anatomical Study of The Blood Supply of The SkinAli HusnainÎncă nu există evaluări
- Heart Sounds:: The Third Sound Is Caused by Vibrations During Passive Filling of The Ventricles With The BloodDocument2 paginiHeart Sounds:: The Third Sound Is Caused by Vibrations During Passive Filling of The Ventricles With The BloodShmuel BarshalomÎncă nu există evaluări
- Ento - Clase 7Document6 paginiEnto - Clase 7KEVIN JOSE GUA�A CATICUAGOÎncă nu există evaluări
- Anatomy of The Heart & Its ValvesDocument9 paginiAnatomy of The Heart & Its ValvesJustine CastilloÎncă nu există evaluări
- Anatomía Del Diafragma NormalDocument7 paginiAnatomía Del Diafragma Normalomar morelosÎncă nu există evaluări
- Jurnal - Christopher Muhamad Zildan Holliday - 215090107111029 - Kelas ADocument17 paginiJurnal - Christopher Muhamad Zildan Holliday - 215090107111029 - Kelas AChristopher Muhamad Zildan HÎncă nu există evaluări
- Respiratory Mechanics and Gas ExchangeDocument14 paginiRespiratory Mechanics and Gas ExchangeNithyaa SathishÎncă nu există evaluări
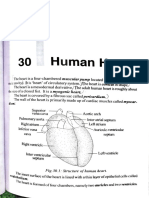
- Chapter 9Document28 paginiChapter 9Jill ZabalaÎncă nu există evaluări
- Supernomal ConductionDocument19 paginiSupernomal ConductionGabriel JoséÎncă nu există evaluări
- Whitaker 2010Document3 paginiWhitaker 2010Gabriel Jose CarvajalÎncă nu există evaluări
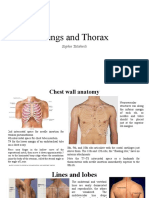
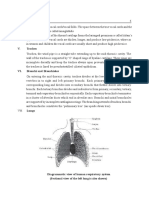
- Lungs and Thorax: Sophie TatishviliDocument31 paginiLungs and Thorax: Sophie TatishviliAstha Tusharbhai PatelÎncă nu există evaluări
- 026 031 PDFDocument6 pagini026 031 PDFStrangersÎncă nu există evaluări
- CH 19-20Document8 paginiCH 19-20yunqiwen2018Încă nu există evaluări
- Anestesia Torax 2Document16 paginiAnestesia Torax 2Oscar Escobedo DamianÎncă nu există evaluări
- Anatomy of The Thoracic Wall, Pulmonary Cavities, and MediastinumDocument26 paginiAnatomy of The Thoracic Wall, Pulmonary Cavities, and Mediastinumearly intervention centre thudiylu trerÎncă nu există evaluări
- Articulo DX Semana 14Document7 paginiArticulo DX Semana 14Valeria Garcia QuijanoÎncă nu există evaluări
- Anatomy of The Dog (186-188)Document4 paginiAnatomy of The Dog (186-188)Natalia Castellanos CoteÎncă nu există evaluări
- THORAX IN FUNNEL - Jhennyfertorres, Yeraldin, Mariaisabel, Adriana2020Document2 paginiTHORAX IN FUNNEL - Jhennyfertorres, Yeraldin, Mariaisabel, Adriana2020Paloma TorresÎncă nu există evaluări
- Lungs&Bronchopulmonary SegmentDocument10 paginiLungs&Bronchopulmonary Segmentshuvam sanatiÎncă nu există evaluări
- Thorax 1Document10 paginiThorax 1Khushi ShahÎncă nu există evaluări
- Tugas DR DewiDocument2 paginiTugas DR DewiAmalia IsnainiÎncă nu există evaluări
- Efecto de MasaDocument7 paginiEfecto de MasaJorge RojasÎncă nu există evaluări
- Blood Supply of Upper LimbDocument36 paginiBlood Supply of Upper Limbteklay100% (2)
- CHAPTER 57 Anatomy and Surgical Access of The MediastinumDocument6 paginiCHAPTER 57 Anatomy and Surgical Access of The MediastinumAbeÎncă nu există evaluări
- Miología Apendicular Del Procyon Cancrivorus (Windle, 1888)Document9 paginiMiología Apendicular Del Procyon Cancrivorus (Windle, 1888)Sebastian ChungaÎncă nu există evaluări
- Almeida Et AlDocument9 paginiAlmeida Et AlGhaniyyatul KhudriÎncă nu există evaluări
- JAssocChestPhysicians1232-7664861 211728Document6 paginiJAssocChestPhysicians1232-7664861 211728Anonymous YYWsfmocYÎncă nu există evaluări
- CVS FullDocument32 paginiCVS FullLianne PalinsadÎncă nu există evaluări
- Advances in Arrhythmia and Electrophysiology: Fascicular Ventricular ArrhythmiasDocument14 paginiAdvances in Arrhythmia and Electrophysiology: Fascicular Ventricular ArrhythmiasMarceli AnjosÎncă nu există evaluări
- The Posterior Fossa VeinsDocument24 paginiThe Posterior Fossa Veinsbodeadumitru9261100% (2)
- Chest X-RayDocument31 paginiChest X-RayPraneetha NouduriÎncă nu există evaluări
- Doppler Venoso de Extremidad SuperiorDocument10 paginiDoppler Venoso de Extremidad SuperiorROBERTO HERNANDEZÎncă nu există evaluări
- Case 2: David Marsden Case Summary: Ostia TrigonesDocument15 paginiCase 2: David Marsden Case Summary: Ostia TrigonesHuiLinn LeeÎncă nu există evaluări
- Breathing For Singing The Anatomy of RespirationDocument11 paginiBreathing For Singing The Anatomy of Respirationbotmy banmeÎncă nu există evaluări
- Mammalian Heart PDFDocument5 paginiMammalian Heart PDFKhaled TurkÎncă nu există evaluări
- V. Trachea: Zoology-IIDocument22 paginiV. Trachea: Zoology-IIVenkatesh JoguÎncă nu există evaluări
- Scan Dec 12, 2022Document6 paginiScan Dec 12, 2022Saktheeswaran VÎncă nu există evaluări
- 13-Vascular of Upper LimbDocument33 pagini13-Vascular of Upper Limbchochojk36Încă nu există evaluări
- The Heart and CirculationDocument7 paginiThe Heart and CirculationsaframikdarhafisÎncă nu există evaluări
- Peckhamia 30Document15 paginiPeckhamia 30api-3703742100% (1)
- Peckhamia 71Document2 paginiPeckhamia 71api-3703742100% (1)
- Peckhamia 65Document4 paginiPeckhamia 65api-3703742Încă nu există evaluări
- Peckhamia 72Document1 paginăPeckhamia 72api-3703742Încă nu există evaluări
- Peckhamia 68Document3 paginiPeckhamia 68api-3703742Încă nu există evaluări
- Peckhamia 64Document2 paginiPeckhamia 64api-3703742Încă nu există evaluări
- Peckhamia 69Document27 paginiPeckhamia 69api-3703742Încă nu există evaluări
- Peckhamia 36Document1 paginăPeckhamia 36api-3703742Încă nu există evaluări
- Peckhamia 70Document35 paginiPeckhamia 70api-3703742Încă nu există evaluări
- Peckhamia 54Document1 paginăPeckhamia 54api-3703742Încă nu există evaluări
- Peckhamia 50Document1 paginăPeckhamia 50api-3703742Încă nu există evaluări
- Peckhamia 53Document1 paginăPeckhamia 53api-3703742Încă nu există evaluări
- Peckhamia 58Document1 paginăPeckhamia 58api-3703742Încă nu există evaluări
- Peckhamia 37Document1 paginăPeckhamia 37api-3703742Încă nu există evaluări
- Peckhamia 43Document1 paginăPeckhamia 43api-3703742Încă nu există evaluări
- Peckhamia 22Document1 paginăPeckhamia 22api-3703742Încă nu există evaluări
- Peckhamia 23Document1 paginăPeckhamia 23api-3703742Încă nu există evaluări
- Peckhamia 67Document1 paginăPeckhamia 67api-3703742Încă nu există evaluări
- Peckhamia 17Document1 paginăPeckhamia 17api-3703742Încă nu există evaluări
- Peckhamia 61Document4 paginiPeckhamia 61api-3703742Încă nu există evaluări
- Peckhamia 66Document4 paginiPeckhamia 66api-3703742Încă nu există evaluări
- Peckhamia 10Document1 paginăPeckhamia 10api-3703742Încă nu există evaluări
- Peckhamia 63Document1 paginăPeckhamia 63api-3703742Încă nu există evaluări
- Peckhamia 52Document3 paginiPeckhamia 52api-3703742Încă nu există evaluări
- Peckhamia 51Document1 paginăPeckhamia 51api-3703742Încă nu există evaluări
- Peckhamia 59Document2 paginiPeckhamia 59api-3703742Încă nu există evaluări
- Peckhamia 60Document6 paginiPeckhamia 60api-3703742Încă nu există evaluări
- Peckhamia 56Document1 paginăPeckhamia 56api-3703742Încă nu există evaluări
- Peckhamia 57Document1 paginăPeckhamia 57api-3703742Încă nu există evaluări
- Peckhamia 49Document3 paginiPeckhamia 49api-3703742Încă nu există evaluări
- BCM 202 Quality AssuranceDocument18 paginiBCM 202 Quality AssuranceTeeÎncă nu există evaluări
- EHBADocument14 paginiEHBAJennifer DixonÎncă nu există evaluări
- Breathing ExercisesDocument16 paginiBreathing ExercisesSubramanya Seshagiri100% (1)
- + +Over+40+Ab+Solution+PDF eBook,+Shaun+HadsallDocument65 pagini+ +Over+40+Ab+Solution+PDF eBook,+Shaun+HadsallSherif Abdel Hamid Fakhry100% (2)
- Unit 2Document8 paginiUnit 2Mason BruceÎncă nu există evaluări
- Apical Third and Its SignificanceDocument20 paginiApical Third and Its SignificanceBharath Ratnakaram100% (1)
- 11 класс 4 модульDocument4 pagini11 класс 4 модульYuriÎncă nu există evaluări
- Soal Kelas 9 Agustus FixDocument2 paginiSoal Kelas 9 Agustus FixYuli KurniatiÎncă nu există evaluări
- Effects of Early Mobilisation in Patients After Cardiac PDFDocument33 paginiEffects of Early Mobilisation in Patients After Cardiac PDFImran TarmiziÎncă nu există evaluări
- Bio Lab # 6Document5 paginiBio Lab # 6Cris SanchEzÎncă nu există evaluări
- RBC CountsDocument16 paginiRBC Countsalmastar officeÎncă nu există evaluări
- Chapter 2 BotanyDocument6 paginiChapter 2 BotanyLebanan Aprille MarieÎncă nu există evaluări
- Module: Lie Detection Techniques (3/1) : at The End of This Chapter The Student's Should Be Able To UnderstandDocument84 paginiModule: Lie Detection Techniques (3/1) : at The End of This Chapter The Student's Should Be Able To Understandjonas ventolinaÎncă nu există evaluări
- Wax and EstersDocument7 paginiWax and EstersPamela Mae UcabÎncă nu există evaluări
- Sexual JealousyDocument11 paginiSexual JealousyLip Stick100% (1)
- ELS Quarter2 OrganSystemsDocument28 paginiELS Quarter2 OrganSystemsJustineÎncă nu există evaluări
- Farmakologi Cns Stimulants DrugsDocument15 paginiFarmakologi Cns Stimulants DrugsRifqiÎncă nu există evaluări
- JCB 201903033Document20 paginiJCB 201903033Purine ChengÎncă nu există evaluări
- Blood Everywhere: A Case Study in BloodDocument2 paginiBlood Everywhere: A Case Study in BloodAbdullahÎncă nu există evaluări
- Med MapsDocument117 paginiMed MapsDukeÎncă nu există evaluări
- Biology 1Document15 paginiBiology 1maielsherif2020Încă nu există evaluări
- Similarities Between Cellular Respiration and PhotosynthesisDocument4 paginiSimilarities Between Cellular Respiration and PhotosynthesisShelline OyooÎncă nu există evaluări
- NCPPPPDocument6 paginiNCPPPPIvan Liquiran AvenadoÎncă nu există evaluări
- ACM 603 Operation and Service ManualDocument26 paginiACM 603 Operation and Service Manualhluevano100% (1)
- CBC ReportsDocument2 paginiCBC ReportsIbrar Hussain100% (1)
- Pharmacology-II NIRALIDocument244 paginiPharmacology-II NIRALIRock Patel93% (29)
- CaPNaK Chart Intro CurrentDocument2 paginiCaPNaK Chart Intro CurrentIrina Elena50% (2)
- 10phedu105 ThesisDocument217 pagini10phedu105 Thesisshruti100% (1)
- In Vivo: Xanthine Oxidase Inhibition StudiesDocument16 paginiIn Vivo: Xanthine Oxidase Inhibition StudiesHartono TanambellÎncă nu există evaluări
- Myocardial InfarctionDocument20 paginiMyocardial InfarctionRio Ramon HilarioÎncă nu există evaluări
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDe la EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceEvaluare: 4.5 din 5 stele4.5/5 (517)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDe la EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisEvaluare: 3.5 din 5 stele3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDe la EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityEvaluare: 4 din 5 stele4/5 (4)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDe la EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsEvaluare: 4.5 din 5 stele4.5/5 (6)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDe la Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessEvaluare: 4 din 5 stele4/5 (33)
- Gut: the new and revised Sunday Times bestsellerDe la EverandGut: the new and revised Sunday Times bestsellerEvaluare: 4 din 5 stele4/5 (393)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedDe la EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedEvaluare: 4 din 5 stele4/5 (11)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeDe la EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeÎncă nu există evaluări
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)De la EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Evaluare: 4 din 5 stele4/5 (411)
- Tales from Both Sides of the Brain: A Life in NeuroscienceDe la EverandTales from Both Sides of the Brain: A Life in NeuroscienceEvaluare: 3 din 5 stele3/5 (18)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionDe la EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionEvaluare: 4 din 5 stele4/5 (812)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesDe la EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesEvaluare: 4.5 din 5 stele4.5/5 (397)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomDe la EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomEvaluare: 4 din 5 stele4/5 (216)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseDe la EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseEvaluare: 4.5 din 5 stele4.5/5 (52)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorDe la EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorÎncă nu există evaluări
- Who's in Charge?: Free Will and the Science of the BrainDe la EverandWho's in Charge?: Free Will and the Science of the BrainEvaluare: 4 din 5 stele4/5 (65)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouDe la EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouEvaluare: 4.5 din 5 stele4.5/5 (62)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperDe la EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperEvaluare: 4.5 din 5 stele4.5/5 (15)
- Seven and a Half Lessons About the BrainDe la EverandSeven and a Half Lessons About the BrainEvaluare: 4 din 5 stele4/5 (109)
- Why We Sleep: Unlocking the Power of Sleep and DreamsDe la EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsEvaluare: 4.5 din 5 stele4.5/5 (2083)
- Lymph & Longevity: The Untapped Secret to HealthDe la EverandLymph & Longevity: The Untapped Secret to HealthEvaluare: 4.5 din 5 stele4.5/5 (13)
- Human: The Science Behind What Makes Your Brain UniqueDe la EverandHuman: The Science Behind What Makes Your Brain UniqueEvaluare: 3.5 din 5 stele3.5/5 (38)
- Good Without God: What a Billion Nonreligious People Do BelieveDe la EverandGood Without God: What a Billion Nonreligious People Do BelieveEvaluare: 4 din 5 stele4/5 (66)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldDe la EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldEvaluare: 4 din 5 stele4/5 (595)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemDe la EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemEvaluare: 4.5 din 5 stele4.5/5 (115)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindDe la EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindEvaluare: 4.5 din 5 stele4.5/5 (93)