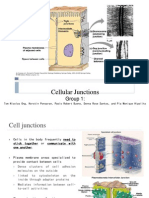
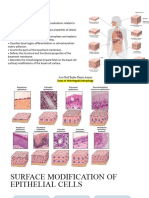
S-ar putea să vă placă și
- From Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.De la EverandFrom Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.Încă nu există evaluări
- Cell Cytoskeleton, Cell JunctionsDocument46 paginiCell Cytoskeleton, Cell JunctionsAhmed khanÎncă nu există evaluări
- Cell JunctionsDocument40 paginiCell JunctionsAhmad MursyidÎncă nu există evaluări
- 05lect Adhesion Summary2017Document5 pagini05lect Adhesion Summary2017Dan ChoiÎncă nu există evaluări
- The CytoskeletonDocument63 paginiThe CytoskeletonAmara TargaryenÎncă nu există evaluări
- BIO130 Sec2 - Lec9and10 - 1pptDocument47 paginiBIO130 Sec2 - Lec9and10 - 1pptlemonpartymanÎncă nu există evaluări
- 19) Integrating Cells Into TissuesDocument37 pagini19) Integrating Cells Into TissuesRÎncă nu există evaluări
- Cell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemDocument70 paginiCell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemGovinda BiswasÎncă nu există evaluări
- BBS1 HS K4 Cell Junction Intercellular CommunicationDocument26 paginiBBS1 HS K4 Cell Junction Intercellular CommunicationVania IlmiÎncă nu există evaluări
- Bloc 8 La Cel Lula I L'entornDocument7 paginiBloc 8 La Cel Lula I L'entornLaia Baus RevueltaÎncă nu există evaluări
- CellDocument47 paginiCellAnami NizamÎncă nu există evaluări
- Cell Junctions and Cell AdhesionDocument31 paginiCell Junctions and Cell AdhesionCitrawati Dyah Kencono WunguÎncă nu există evaluări
- C & M Bio CytoskeletonDocument40 paginiC & M Bio CytoskeletonA PÎncă nu există evaluări
- Cell-Cell Junctions: Bhavtosh A. Kikani, PHD, Icar-NetDocument16 paginiCell-Cell Junctions: Bhavtosh A. Kikani, PHD, Icar-NetB KikaniÎncă nu există evaluări
- of Cell Adhesion and Cell JunctionsDocument50 paginiof Cell Adhesion and Cell Junctionsayad ghawiÎncă nu există evaluări
- Bio2000 Notes-2004-2Document21 paginiBio2000 Notes-2004-2api-3700537Încă nu există evaluări
- Cell JunctionsDocument30 paginiCell JunctionsNavodit GoelÎncă nu există evaluări
- Mammalian Histology AssignmentDocument9 paginiMammalian Histology AssignmentSana Sultana100% (1)
- Cell JunctionsDocument34 paginiCell JunctionsDrAmit Gaba Mds100% (1)
- Bookmark This Page: Cadherins Mucin-Like Cams IntegrinsDocument38 paginiBookmark This Page: Cadherins Mucin-Like Cams IntegrinsAndreea SpiridonÎncă nu există evaluări
- Cell Junction, Adhesion and Extracellular MatrixDocument14 paginiCell Junction, Adhesion and Extracellular Matrixseenu mohapatraÎncă nu există evaluări
- Cell JunctionsDocument24 paginiCell JunctionsNajlaÎncă nu există evaluări
- Cell JunctionsDocument56 paginiCell JunctionsDrGagandeep Singh100% (3)
- Epithelium Surface SpecializationsDocument41 paginiEpithelium Surface SpecializationsMitzel SapaloÎncă nu există evaluări
- Cellular JunctionsDocument25 paginiCellular JunctionsDiana Grace Pile0% (1)
- Cell JunctionsDocument1 paginăCell JunctionsJoshua Mari De OcampoÎncă nu există evaluări
- Cell Bio 3 Cell Junctions 2021 PDFDocument38 paginiCell Bio 3 Cell Junctions 2021 PDFaboodmr8nblahÎncă nu există evaluări
- Introduction To Histology and Epithelium TissueDocument48 paginiIntroduction To Histology and Epithelium TissuejabirÎncă nu există evaluări
- JI UNIT 2 Complete Notes - 7-8Document2 paginiJI UNIT 2 Complete Notes - 7-8Shubham PawadeÎncă nu există evaluări
- Junctions - 2008 ArisDocument34 paginiJunctions - 2008 ArisAmardeep SinghÎncă nu există evaluări
- Cell JunctionDocument63 paginiCell JunctionsphuyalÎncă nu există evaluări
- Extracellular Matrix and CytoskeletonDocument23 paginiExtracellular Matrix and CytoskeletonjoyyeesufÎncă nu există evaluări
- 2004 4 JW CellJunctions ELShhhhhhDocument7 pagini2004 4 JW CellJunctions ELShhhhhhhfqjhrpjncÎncă nu există evaluări
- Plant Cells Structure of The Plant CellsDocument10 paginiPlant Cells Structure of The Plant Cellsstudent10100Încă nu există evaluări
- Cell To Cell ContactDocument6 paginiCell To Cell ContactHarini BalasubramanianÎncă nu există evaluări
- Unit 15 Cell Junction DhanDocument18 paginiUnit 15 Cell Junction DhanSarah PavuÎncă nu există evaluări
- CytoskeletonDocument86 paginiCytoskeletonAbid Al RezaÎncă nu există evaluări
- Cell-to-Cell and Cell-to-Matrix Adhesions: Fibronectin?Document46 paginiCell-to-Cell and Cell-to-Matrix Adhesions: Fibronectin?MinarsihÎncă nu există evaluări
- Cell Junctions, Cell-Cell AdhesionsDocument36 paginiCell Junctions, Cell-Cell AdhesionsRizky Nur IndahÎncă nu există evaluări
- JunctionsDocument96 paginiJunctionsAnyone Can CookÎncă nu există evaluări
- Cytoskeleton: Jetro P. MagnayonDocument66 paginiCytoskeleton: Jetro P. MagnayonChristian Santiago100% (1)
- BM 1 1 CytoskeletonDocument24 paginiBM 1 1 CytoskeletonSanthoshi Sadhanaa SankarÎncă nu există evaluări
- Molecular Bioeng-2: Mrs Daramola BellstechDocument22 paginiMolecular Bioeng-2: Mrs Daramola BellstechSamuel EkpoÎncă nu există evaluări
- Cytoskeleton, Junctions, Fibroblast and ECMDocument38 paginiCytoskeleton, Junctions, Fibroblast and ECMManoj HumagainÎncă nu există evaluări
- 2 Basal Lateral Apical Modifications and Connective TissueDocument84 pagini2 Basal Lateral Apical Modifications and Connective TissueDeniz asmazÎncă nu există evaluări
- Lewin's Cells 3rd Ed 2015 Ranit-855-914Document60 paginiLewin's Cells 3rd Ed 2015 Ranit-855-914Muhammad Ikram RabbaniÎncă nu există evaluări
- Cell Adhesion and Cell Adhesion MoleculesDocument7 paginiCell Adhesion and Cell Adhesion MoleculesLemon GundersonÎncă nu există evaluări
- 2structure and Functions of Cell MembraneDocument19 pagini2structure and Functions of Cell MembraneedriansamaÎncă nu există evaluări
- Cell JunctionsDocument18 paginiCell Junctionsrevathikarri065Încă nu există evaluări
- Cellular Nanomechanics 2Document27 paginiCellular Nanomechanics 2Ahmed MotasimÎncă nu există evaluări
- Cells As Units of LifeDocument65 paginiCells As Units of LifeBenjamin CoÎncă nu există evaluări
- Cytoskeleton DADocument35 paginiCytoskeleton DAAnonymous NVBCqKWHÎncă nu există evaluări
- February 27, 2012 - Chapter 7: Inside The CellDocument4 paginiFebruary 27, 2012 - Chapter 7: Inside The CellVincent Troy SantosÎncă nu există evaluări
- 2016 10 18 The Extracellular Matrix (ECM)Document45 pagini2016 10 18 The Extracellular Matrix (ECM)sennaavia12100% (1)
- Partoza, J WS Cel - MolDocument6 paginiPartoza, J WS Cel - MolJenny PartozaÎncă nu există evaluări
- EEU2020 Adfinex HDocument23 paginiEEU2020 Adfinex HZainab Jamal SiddiquiÎncă nu există evaluări
- Cell Adhesion Molecules 300107Document32 paginiCell Adhesion Molecules 300107V Lee 'Nozhat'100% (1)
- 4 - Cell JunctionsDocument25 pagini4 - Cell JunctionsKhdhir M. Salem100% (1)
- Cytoskeleton Motor Proteins and ECM - SCDocument77 paginiCytoskeleton Motor Proteins and ECM - SCAlice RizzoÎncă nu există evaluări
- Physiological Processes Where Cell Adhesion Is ImportantDocument42 paginiPhysiological Processes Where Cell Adhesion Is ImportantJT92Încă nu există evaluări
- Lecture 26 NotesDocument3 paginiLecture 26 Notesapi-3721438Încă nu există evaluări
- Lecture 25 NotesDocument4 paginiLecture 25 Notesapi-3721438Încă nu există evaluări
- Lecture 28 NotesDocument6 paginiLecture 28 Notesapi-3721438100% (1)
- Lectures 1-7 CH15 NotesDocument29 paginiLectures 1-7 CH15 Notesapi-3721438Încă nu există evaluări
- Lectures 8-10 NotesDocument15 paginiLectures 8-10 Notesapi-3721438Încă nu există evaluări
- Inducible GenesDocument6 paginiInducible Genesapi-3721438Încă nu există evaluări
- GeneDocument8 paginiGeneapi-3721438100% (1)
- AT10 Meat Tech 1Document20 paginiAT10 Meat Tech 1Reubal Jr Orquin Reynaldo100% (1)
- Caddy Arc 251i Service ManualDocument52 paginiCaddy Arc 251i Service Manualrikycien100% (8)
- Encapsulation of Objects and Methods in C++Document46 paginiEncapsulation of Objects and Methods in C++Scott StanleyÎncă nu există evaluări
- Space Saving, Tight AccessibilityDocument4 paginiSpace Saving, Tight AccessibilityTran HuyÎncă nu există evaluări
- Conjunctions in SentencesDocument8 paginiConjunctions in SentencesPunitha PoppyÎncă nu există evaluări
- One Foot in The Grave - Copy For PlayersDocument76 paginiOne Foot in The Grave - Copy For Playerssveni meierÎncă nu există evaluări
- EPSS 627: DescriptionDocument2 paginiEPSS 627: DescriptionudayakumartÎncă nu există evaluări
- A Project Report On "A Comparative Study Between Hero Honda Splendor+ and Its Competitors To Increase The Market Share in MUDHOL RegionDocument70 paginiA Project Report On "A Comparative Study Between Hero Honda Splendor+ and Its Competitors To Increase The Market Share in MUDHOL RegionBabasab Patil (Karrisatte)Încă nu există evaluări
- CCTV Guidelines - Commission Letter Dated 27.08.2022Document2 paginiCCTV Guidelines - Commission Letter Dated 27.08.2022Sumeet TripathiÎncă nu există evaluări
- Maintenance Performance ToolboxDocument6 paginiMaintenance Performance ToolboxMagda ScrobotaÎncă nu există evaluări
- Fh84fr6ht GBR EngDocument6 paginiFh84fr6ht GBR EngEsmir ŠkreboÎncă nu există evaluări
- Computerized Flat Knitting Machine Computerized Flat Knitting Machine Computerized Flat Knitting Machine Computerized Flat Knitting MachineDocument61 paginiComputerized Flat Knitting Machine Computerized Flat Knitting Machine Computerized Flat Knitting Machine Computerized Flat Knitting MachineAmira's ClothesÎncă nu există evaluări
- Genie PDFDocument264 paginiGenie PDFjohanaÎncă nu există evaluări
- NDT HandBook Volume 10 (NDT Overview)Document600 paginiNDT HandBook Volume 10 (NDT Overview)mahesh95% (19)
- Intraoperative RecordDocument2 paginiIntraoperative Recordademaala06100% (1)
- De On Tap So 4-6Document8 paginiDe On Tap So 4-6Quy DoÎncă nu există evaluări
- What Is An EcosystemDocument42 paginiWhat Is An Ecosystemjoniel05Încă nu există evaluări
- Travelstart Ticket (ZA10477979) PDFDocument2 paginiTravelstart Ticket (ZA10477979) PDFMatthew PretoriusÎncă nu există evaluări
- POLYTHEOREMSDocument32 paginiPOLYTHEOREMSYen LeeÎncă nu există evaluări
- Heisenberg, "Über Den Anschaulichen Inhalt Der Quantentheoretischen Kinematik Und Mechanik"Document16 paginiHeisenberg, "Über Den Anschaulichen Inhalt Der Quantentheoretischen Kinematik Und Mechanik"Benjamin Crowell0% (1)
- Letter Writing: An Informative Powerpoint About LetterDocument11 paginiLetter Writing: An Informative Powerpoint About LetterMalik KamranÎncă nu există evaluări
- Development of A Small Solar Thermal PowDocument10 paginiDevelopment of A Small Solar Thermal Powעקיבא אסÎncă nu există evaluări
- My Activities in Module 2Document7 paginiMy Activities in Module 2Devine Gabat100% (6)
- Advanced Statistical Approaches To Quality: INSE 6220 - Week 4Document44 paginiAdvanced Statistical Approaches To Quality: INSE 6220 - Week 4picalaÎncă nu există evaluări
- 24 Inch MonitorDocument10 pagini24 Inch MonitorMihir SaveÎncă nu există evaluări
- CSR Report On Tata SteelDocument72 paginiCSR Report On Tata SteelJagadish Sahu100% (1)
- Image Hosting SitesDocument16 paginiImage Hosting SitesstudentÎncă nu există evaluări
- Appendix 3 COT RPMS For T I III SY 2020 2021 in The Time of COVID 19Document12 paginiAppendix 3 COT RPMS For T I III SY 2020 2021 in The Time of COVID 19Marjun PachecoÎncă nu există evaluări
- Ajsl DecisionMakingModel4RoRoDocument11 paginiAjsl DecisionMakingModel4RoRolesta putriÎncă nu există evaluări
- 12 Constructor and DistructorDocument15 pagini12 Constructor and DistructorJatin BhasinÎncă nu există evaluări
- Why We Die: The New Science of Aging and the Quest for ImmortalityDe la EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityEvaluare: 4 din 5 stele4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDe la EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisEvaluare: 3.5 din 5 stele3.5/5 (2)
- Gut: the new and revised Sunday Times bestsellerDe la EverandGut: the new and revised Sunday Times bestsellerEvaluare: 4 din 5 stele4/5 (392)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesDe la EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesEvaluare: 4.5 din 5 stele4.5/5 (397)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeDe la EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeÎncă nu există evaluări
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDe la EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsEvaluare: 4.5 din 5 stele4.5/5 (6)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDe la EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceEvaluare: 4.5 din 5 stele4.5/5 (517)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)De la EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Evaluare: 4 din 5 stele4/5 (378)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDe la Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessEvaluare: 4 din 5 stele4/5 (33)
- Tales from Both Sides of the Brain: A Life in NeuroscienceDe la EverandTales from Both Sides of the Brain: A Life in NeuroscienceEvaluare: 3 din 5 stele3/5 (18)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionDe la EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionEvaluare: 4 din 5 stele4/5 (812)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperDe la EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperEvaluare: 4.5 din 5 stele4.5/5 (15)
- Good Without God: What a Billion Nonreligious People Do BelieveDe la EverandGood Without God: What a Billion Nonreligious People Do BelieveEvaluare: 4 din 5 stele4/5 (66)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouDe la EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouEvaluare: 4.5 din 5 stele4.5/5 (62)
- Seven and a Half Lessons About the BrainDe la EverandSeven and a Half Lessons About the BrainEvaluare: 4 din 5 stele4/5 (109)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedDe la EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedEvaluare: 4 din 5 stele4/5 (11)
- Who's in Charge?: Free Will and the Science of the BrainDe la EverandWho's in Charge?: Free Will and the Science of the BrainEvaluare: 4 din 5 stele4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorDe la EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorÎncă nu există evaluări
- Human: The Science Behind What Makes Your Brain UniqueDe la EverandHuman: The Science Behind What Makes Your Brain UniqueEvaluare: 3.5 din 5 stele3.5/5 (38)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldDe la EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldEvaluare: 4 din 5 stele4/5 (595)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindDe la EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindEvaluare: 4.5 din 5 stele4.5/5 (93)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemDe la EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemEvaluare: 4.5 din 5 stele4.5/5 (115)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldDe la EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldEvaluare: 4.5 din 5 stele4.5/5 (18)
- Why We Sleep: Unlocking the Power of Sleep and DreamsDe la EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsEvaluare: 4.5 din 5 stele4.5/5 (2083)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineDe la EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineEvaluare: 4 din 5 stele4/5 (17)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomDe la EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomEvaluare: 4 din 5 stele4/5 (216)