S-ar putea să vă placă și
- Neurofeedback: Emotive EEG Epoch Is A Research Grade DeviceDocument19 paginiNeurofeedback: Emotive EEG Epoch Is A Research Grade Deviceavalon_moonÎncă nu există evaluări
- Fuel Economy of Road VehiclesDocument50 paginiFuel Economy of Road VehiclesdressfeetÎncă nu există evaluări
- Mindfulness Meditation and Cognitive Behavioural Therapy For TinnitusDocument3 paginiMindfulness Meditation and Cognitive Behavioural Therapy For Tinnitusavalon_moonÎncă nu există evaluări
- Positive Psychology and Adolescent Mental HealthDocument16 paginiPositive Psychology and Adolescent Mental Healthavalon_moonÎncă nu există evaluări
- Carlos Herraiz - HYPERACUSISDocument45 paginiCarlos Herraiz - HYPERACUSISavalon_moonÎncă nu există evaluări
- Noise-Induced Hearing Loss and TinnitusDocument167 paginiNoise-Induced Hearing Loss and Tinnitusavalon_moonÎncă nu există evaluări
- Limbic System Our Emotional CenterDocument27 paginiLimbic System Our Emotional Centeravalon_moonÎncă nu există evaluări
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (400)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (74)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Behavioural Observation AudiometryDocument4 paginiBehavioural Observation AudiometryLubaba lubinpÎncă nu există evaluări
- AudiDocument3 paginiAudiFlorygene Kris Disagon0% (1)
- 6 - Calulation of Noise LevelsDocument12 pagini6 - Calulation of Noise Levelsjun005Încă nu există evaluări
- Learning CompetenciesDocument9 paginiLearning CompetenciesVencint Karl Tacardon100% (1)
- EAR MCQs Points Rida NaqviDocument6 paginiEAR MCQs Points Rida NaqviÅli Raza ChaudaryÎncă nu există evaluări
- Operating Instructions MA 42 - Maico Diagnostics PDFDocument28 paginiOperating Instructions MA 42 - Maico Diagnostics PDFJuan PáezÎncă nu există evaluări
- Technical Brief :recommendedDocument1 paginăTechnical Brief :recommendedglens_nwa100% (2)
- Common ENT Conditions PwPT2008Document75 paginiCommon ENT Conditions PwPT2008Garlic BreadÎncă nu există evaluări
- Lecture 3 - Communication ApproachesDocument89 paginiLecture 3 - Communication Approachesapi-349133705Încă nu există evaluări
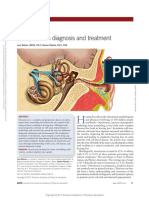
- Otosclerosis An Update On Diagnosis and Treatment.3Document6 paginiOtosclerosis An Update On Diagnosis and Treatment.3andiÎncă nu există evaluări
- Physio HearingDocument56 paginiPhysio Hearing65 Nidheesh Kumar PrabakamoorthyÎncă nu există evaluări
- DM-PH&SD-P4-TG10 - (Guidelines For Personal Protective Equipment-Hearing Protection) PDFDocument3 paginiDM-PH&SD-P4-TG10 - (Guidelines For Personal Protective Equipment-Hearing Protection) PDFdemie figueroaÎncă nu există evaluări
- Grade 3 2nd Quarter Unit 3 Lesson 1 Sense Organs and How To Care For ThemDocument89 paginiGrade 3 2nd Quarter Unit 3 Lesson 1 Sense Organs and How To Care For ThemRaul Rivera SantosÎncă nu există evaluări
- Hearing Loss (Badhirya) and Its Ayurvedic Management: A Case StudyDocument9 paginiHearing Loss (Badhirya) and Its Ayurvedic Management: A Case Studyshweta kotwal100% (1)
- ENCOREPRO 510/520: Leave A Lasting ImpressionDocument2 paginiENCOREPRO 510/520: Leave A Lasting Impressionraoultrifan7560Încă nu există evaluări
- Amazing Mind TricksDocument11 paginiAmazing Mind Tricksdarl1Încă nu există evaluări
- Magic FrequenciesDocument6 paginiMagic FrequenciesWagner SilvaÎncă nu există evaluări
- June Wieder - Song of The Spine (2004)Document128 paginiJune Wieder - Song of The Spine (2004)constelacionesparalavidaÎncă nu există evaluări
- 2002 (ProAudio) Sound Engineering - Advice On Mixing - Bill Gibson PDFDocument39 pagini2002 (ProAudio) Sound Engineering - Advice On Mixing - Bill Gibson PDFMauricio Rodriguez WylerÎncă nu există evaluări
- Dpa 4099 Guide PDFDocument4 paginiDpa 4099 Guide PDFEmiliana SilvaÎncă nu există evaluări
- A Movie Review Mr. Holland's OpusDocument6 paginiA Movie Review Mr. Holland's OpusAnne MarielÎncă nu există evaluări
- Strahlenfolter - Ultrasonic DevicesDocument5 paginiStrahlenfolter - Ultrasonic Devicesstop-nwo-nazi-folter100% (1)
- DDocument4 paginiDPraveenaslp EddulaÎncă nu există evaluări
- 4 5877209711425619242Document92 pagini4 5877209711425619242Thadei MrumaÎncă nu există evaluări
- Things I Learned in Module 5: By: Abdiel CortesDocument5 paginiThings I Learned in Module 5: By: Abdiel CortesABDIEL CORTESÎncă nu există evaluări
- Acoustics: Acoustics Is The Interdisciplinary Science That Deals With The Study of AllDocument3 paginiAcoustics: Acoustics Is The Interdisciplinary Science That Deals With The Study of AllharshalÎncă nu există evaluări
- What Does A Piano Tuner Do When He Goes Deaf PDFDocument4 paginiWhat Does A Piano Tuner Do When He Goes Deaf PDFSpencer GrahamÎncă nu există evaluări
- Curricula Vitate WitulskiDocument4 paginiCurricula Vitate Witulskiapi-247968027Încă nu există evaluări
- Tuning Fork TestsDocument3 paginiTuning Fork TestsClarissa YudakusumaÎncă nu există evaluări