S-ar putea să vă placă și
- Sudden Cardic ArrestDocument17 paginiSudden Cardic ArrestPiet Pogen PoelÎncă nu există evaluări
- Hyper Nat Remi ADocument2 paginiHyper Nat Remi APiet Pogen PoelÎncă nu există evaluări
- Dehydration ProcessDocument7 paginiDehydration ProcessSunil BarnwalÎncă nu există evaluări
- Airway Management - Supraglottic Airway PDFDocument4 paginiAirway Management - Supraglottic Airway PDFPiet Pogen PoelÎncă nu există evaluări
- Basic (Level IV) Trauma Facility CriteriaDocument6 paginiBasic (Level IV) Trauma Facility CriteriaPiet Pogen PoelÎncă nu există evaluări
- Airway Management - Supraglottic Airway PDFDocument4 paginiAirway Management - Supraglottic Airway PDFPiet Pogen PoelÎncă nu există evaluări
- Seeking Patients Informed Consent Ethical ConsiderationDocument18 paginiSeeking Patients Informed Consent Ethical ConsiderationPiet Pogen PoelÎncă nu există evaluări
- Appendix A. THE MEDICAL HISTORY (For New Patient) : Immunizations Injuries Broken Bones/fractures, Sutured LacerationsDocument1 paginăAppendix A. THE MEDICAL HISTORY (For New Patient) : Immunizations Injuries Broken Bones/fractures, Sutured LacerationsPiet Pogen PoelÎncă nu există evaluări
- International Trauma Life Support Takes Trauma Training Online With New Itls Etrauma ProgramDocument2 paginiInternational Trauma Life Support Takes Trauma Training Online With New Itls Etrauma ProgramPiet Pogen PoelÎncă nu există evaluări
- Flow ChartDocument4 paginiFlow ChartPiet Pogen PoelÎncă nu există evaluări
- Flow ChartDocument4 paginiFlow ChartPiet Pogen PoelÎncă nu există evaluări
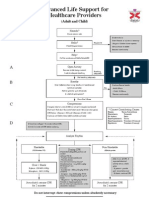
- ALS Adult and ChildDocument1 paginăALS Adult and ChildPiet Pogen PoelÎncă nu există evaluări
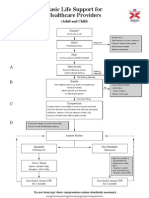
- BLS Adult and ChildDocument1 paginăBLS Adult and ChildPiet Pogen PoelÎncă nu există evaluări
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (120)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- 2010 Hiv Aids GrantsDocument216 pagini2010 Hiv Aids GrantsSherry HeinzÎncă nu există evaluări
- HIV IntroductionDocument27 paginiHIV IntroductionAntónio CantoÎncă nu există evaluări
- Edward C. Klatt - Pathology of HIVAIDS 2018Document420 paginiEdward C. Klatt - Pathology of HIVAIDS 2018BiancaÎncă nu există evaluări
- By Rebecca McmullenDocument10 paginiBy Rebecca McmullenGenaroacÎncă nu există evaluări
- Hivaids - Book ChapterDocument9 paginiHivaids - Book ChapterNnaji OgochukwuÎncă nu există evaluări
- Argumentos en Contra Del Uso de Animales de ExperimentaciónDocument5 paginiArgumentos en Contra Del Uso de Animales de ExperimentaciónJuan David RejónÎncă nu există evaluări
- Hypothetical Possibilities of Development of Non-Hodgkins Lymphoma During Lyme BorreliosisDocument16 paginiHypothetical Possibilities of Development of Non-Hodgkins Lymphoma During Lyme Borreliosisiaset123Încă nu există evaluări
- Humanities Abstracts: "Margaret C. Anderson's Little Review"Document22 paginiHumanities Abstracts: "Margaret C. Anderson's Little Review"franxyz14344Încă nu există evaluări
- Zoo 1100-1Document66 paginiZoo 1100-1BUNDI VINCENTÎncă nu există evaluări
- Klatt 2005 Pathology of AIDSDocument290 paginiKlatt 2005 Pathology of AIDSMaya RustamÎncă nu există evaluări
- Unit 8 Cause & Effect, Comparison & Contrast, and Adverb ClausesDocument14 paginiUnit 8 Cause & Effect, Comparison & Contrast, and Adverb ClausescrrrrÎncă nu există evaluări
- 5 6059713553416323809Document351 pagini5 6059713553416323809Dimas RfÎncă nu există evaluări
- Inform Consent HIVDocument465 paginiInform Consent HIVHartini100% (2)
- Origin of Aids DiseaseDocument13 paginiOrigin of Aids DiseaseRahul SharotriÎncă nu există evaluări
- NIH Public Access: Author ManuscriptDocument33 paginiNIH Public Access: Author ManuscriptPuskesmas RawaloÎncă nu există evaluări
- PM - Materi 1 HIV AIDS RevDocument72 paginiPM - Materi 1 HIV AIDS RevSaly AlatasÎncă nu există evaluări
- Adaptation of HIV-1 To Its Human HostDocument8 paginiAdaptation of HIV-1 To Its Human HostImperialAquillaÎncă nu există evaluări
- No Lie Can Live Forever - Mitchell AguilarDocument265 paginiNo Lie Can Live Forever - Mitchell AguilarNoliecanLiveforever100% (1)
- AIDS Is Not InfectiousDocument18 paginiAIDS Is Not Infectiousbadboy6.868510Încă nu există evaluări
- Hiv/Aids Hiv (Disambiguation) Aids (Computer Virus) : Jump To Navigationjump To SearchDocument16 paginiHiv/Aids Hiv (Disambiguation) Aids (Computer Virus) : Jump To Navigationjump To SearchGita SJÎncă nu există evaluări
- Aids Hiv AssignmentDocument21 paginiAids Hiv AssignmentGeetha Sarika100% (3)
- Origin of HIVDocument18 paginiOrigin of HIVRobert SineÎncă nu există evaluări
- Vaccine Guide - Randall NeustaedterDocument6 paginiVaccine Guide - Randall NeustaedterttreksÎncă nu există evaluări
- Management of The Patient With HIV DeseaseDocument34 paginiManagement of The Patient With HIV DeseaseTeresa MontesÎncă nu există evaluări
- Self Disseminating VaccinesDocument9 paginiSelf Disseminating VaccinesCaesar VranceanuÎncă nu există evaluări
- Enfermedades EmergentesDocument216 paginiEnfermedades EmergentesCaaarolÎncă nu există evaluări
- Ketiga 1 3 EngDocument14 paginiKetiga 1 3 EngAulia putriÎncă nu există evaluări
- Jurnal Hiv Bahasa Inggris 2018Document6 paginiJurnal Hiv Bahasa Inggris 2018talha laayuÎncă nu există evaluări
- ANIMALS Singer MerrillDocument32 paginiANIMALS Singer MerrillBlythe TomÎncă nu există evaluări
- MicrofinalDocument7 paginiMicrofinaljwanaÎncă nu există evaluări