S-ar putea să vă placă și
- Gale Researcher Guide for: Overview of Physiology and NeuropsychologyDe la EverandGale Researcher Guide for: Overview of Physiology and NeuropsychologyÎncă nu există evaluări
- Motor Systems 10 05Document11 paginiMotor Systems 10 05souvik5000100% (1)
- Mind at Rest: How Neuron Structure Evolves in the Sleep Cycle.De la EverandMind at Rest: How Neuron Structure Evolves in the Sleep Cycle.Încă nu există evaluări
- Locomotor System (Text)Document3 paginiLocomotor System (Text)Aknur JakhanshaÎncă nu există evaluări
- Physio - Psych Chapter-8 - Control-Of-Movemnt - Fact-SheetDocument9 paginiPhysio - Psych Chapter-8 - Control-Of-Movemnt - Fact-SheetJustynmae JusainÎncă nu există evaluări
- Motor SystemDocument49 paginiMotor SystemAhaisibwe GordonÎncă nu există evaluări
- Cerebellum in Overall Motor ControlDocument5 paginiCerebellum in Overall Motor Controldncm.alejandroÎncă nu există evaluări
- CORTICAL & BRAINSTEM CONTROL OF MOTOR FUNCTIONSDocument268 paginiCORTICAL & BRAINSTEM CONTROL OF MOTOR FUNCTIONSsaraÎncă nu există evaluări
- Neural Control of LocomotionDocument11 paginiNeural Control of Locomotionphysiovipin86% (7)
- CerebellumDocument51 paginiCerebellumLiya ThaaraÎncă nu există evaluări
- LECTURE On Spinal CordDocument11 paginiLECTURE On Spinal CordElijah KamaniÎncă nu există evaluări
- 3motor TestingDocument74 pagini3motor Testing2013SecBÎncă nu există evaluări
- The Motor System Lecture 1 OverviewDocument7 paginiThe Motor System Lecture 1 OverviewUsman Ali AkbarÎncă nu există evaluări
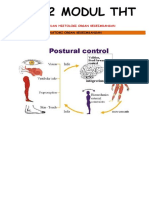
- LBM 2 THTDocument20 paginiLBM 2 THTelly nurkhikmahÎncă nu există evaluări
- L1 Motor Function of CNSDocument48 paginiL1 Motor Function of CNSAhmed JawdetÎncă nu există evaluări
- Spinal ReportDocument28 paginiSpinal ReportJohngie QuilangÎncă nu există evaluări
- Cerebellum: Movement Regulation and Cognitive Functions Course Title DateDocument16 paginiCerebellum: Movement Regulation and Cognitive Functions Course Title DateDanyÎncă nu există evaluări
- Scientific Basis of Body Mapping and Cortical MapsDocument2 paginiScientific Basis of Body Mapping and Cortical MapsDssalamancazÎncă nu există evaluări
- The Cerebelum 2Document53 paginiThe Cerebelum 2Reagan MwesigwaÎncă nu există evaluări
- Cerebellum Essay - Connections and Functions of The CerebellumDocument4 paginiCerebellum Essay - Connections and Functions of The CerebellumscholifyÎncă nu există evaluări
- Physio Psy Module 8Document4 paginiPhysio Psy Module 8Kier Elizalde VistalÎncă nu există evaluări
- Neurophysiology II 2023Document90 paginiNeurophysiology II 2023trsvscpz79Încă nu există evaluări
- Neuro anatomy, motor neuronDocument9 paginiNeuro anatomy, motor neuronMichele Silva PiresÎncă nu există evaluări
- Ectodermal Derivatives of Pig EmbryoDocument7 paginiEctodermal Derivatives of Pig EmbryoChristalie Bea FernandezÎncă nu există evaluări
- ASLI - 20.10.2022NMD-5 Anatomy of Motor ControlDocument40 paginiASLI - 20.10.2022NMD-5 Anatomy of Motor ControlYorgos TheodosiadisÎncă nu există evaluări
- Neuro AnatomyDocument26 paginiNeuro Anatomymeryclave345Încă nu există evaluări
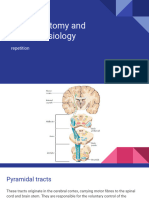
- Physiology of Motor TractsDocument34 paginiPhysiology of Motor TractsNiko MartinÎncă nu există evaluări
- The descending pathways that control motor activityDocument2 paginiThe descending pathways that control motor activityalmastar officeÎncă nu există evaluări
- Cerebellar Function TestDocument37 paginiCerebellar Function TestAdrian Paolo Malamug100% (1)
- SASDocument14 paginiSASNicole Ken AgdanaÎncă nu există evaluări
- Ch. 5Document8 paginiCh. 5Sabrina MaciarielloÎncă nu există evaluări
- Session #22 SAS - AnaPhy (Lab)Document14 paginiSession #22 SAS - AnaPhy (Lab)G IÎncă nu există evaluări
- Jingying T.A. Department of PhysiologyDocument19 paginiJingying T.A. Department of Physiologyapi-19916399Încă nu există evaluări
- PBLDocument3 paginiPBLAbi AuxÎncă nu există evaluări
- Understanding Human Anatomy and Brain FunctionDocument61 paginiUnderstanding Human Anatomy and Brain Functionjustice88_639020831Încă nu există evaluări
- Neurotransmitter Assingment Ashish Singh PDFDocument7 paginiNeurotransmitter Assingment Ashish Singh PDFAshish SinghÎncă nu există evaluări
- Motor Control IIDocument39 paginiMotor Control IILuis Eduardo RibeiroÎncă nu există evaluări
- Skeletal Motor Control HierarchyDocument16 paginiSkeletal Motor Control HierarchyHapy ArdiaÎncă nu există evaluări
- CerebellumDocument16 paginiCerebellumjainilÎncă nu există evaluări
- Systems Neuroscience Nov. 12, 2019Document37 paginiSystems Neuroscience Nov. 12, 2019TrajceÎncă nu există evaluări
- Corticospinal Tract: Functions and Clinical RelevanceDocument4 paginiCorticospinal Tract: Functions and Clinical RelevanceNrs Sani Sule MashiÎncă nu există evaluări
- Somatosensory Pathways: Peripheral Receptors and Central PathwaysDocument28 paginiSomatosensory Pathways: Peripheral Receptors and Central PathwaysJean Pierre Chastre LuzaÎncă nu există evaluări
- Neuroanatomy, Extrapyramidal System: AuthorsDocument7 paginiNeuroanatomy, Extrapyramidal System: AuthorsDiana CastilloÎncă nu există evaluări
- Motor system overview: Upper and lower motor neuronsDocument12 paginiMotor system overview: Upper and lower motor neuronssimi yÎncă nu există evaluări
- Human Nervous System - Movement - BritannicaDocument15 paginiHuman Nervous System - Movement - Britannicasalamy amourÎncă nu există evaluări
- Sensory PathwayDocument31 paginiSensory PathwayAlma BaterinaÎncă nu există evaluări
- Chapter SummaryDocument2 paginiChapter SummaryrogelynlumasagÎncă nu există evaluări
- Motor Control Theories and PhysiologyDocument7 paginiMotor Control Theories and PhysiologyAaminah NawazÎncă nu există evaluări
- Upper Motor Neuron PDFDocument3 paginiUpper Motor Neuron PDFkevin_jawanÎncă nu există evaluări
- Basic Information About NeuropsychologyDocument9 paginiBasic Information About NeuropsychologyYara Bou AbdoÎncă nu există evaluări
- 2.update Sensorik-Motorik NeuropsikiatriDocument56 pagini2.update Sensorik-Motorik NeuropsikiatriumamitÎncă nu există evaluări
- Chapter 10 Body MovementDocument27 paginiChapter 10 Body MovementHarpreet SinghÎncă nu există evaluări
- Nervous System AnswerDocument9 paginiNervous System AnswerVann VelascoÎncă nu există evaluări
- Ballon, Rosemarie E. Chapter 4Document3 paginiBallon, Rosemarie E. Chapter 4Rose Marie BallonÎncă nu există evaluări
- Motor Functions of Spinal CordDocument43 paginiMotor Functions of Spinal CordEugeniu CoretchiÎncă nu există evaluări
- Assesment NeuroDocument15 paginiAssesment Neuroerfan ghadirianÎncă nu există evaluări
- Lo 1 Week 5 NeuroDocument36 paginiLo 1 Week 5 NeuroInna DamaÎncă nu există evaluări
- Hes006 CNS Lab 15 18Document50 paginiHes006 CNS Lab 15 18Joana Elizabeth A. CastroÎncă nu există evaluări
- MBBC 2018 Notes 1Document14 paginiMBBC 2018 Notes 1Dimitri KhooÎncă nu există evaluări
- Exploring the Automatic Responses of ReflexesDocument2 paginiExploring the Automatic Responses of Reflexesmohitnet1327Încă nu există evaluări
- Mandibular FracturesDocument70 paginiMandibular FracturesPatterson MachariaÎncă nu există evaluări
- Developmental Disorders of TeethDocument64 paginiDevelopmental Disorders of TeethPatterson MachariaÎncă nu există evaluări
- Biology of Tooth MovementDocument5 paginiBiology of Tooth MovementPatterson Macharia100% (1)
- Limbic System (Behaviour and Emotion)Document19 paginiLimbic System (Behaviour and Emotion)Patterson MachariaÎncă nu există evaluări
- Development of The MaxillaDocument7 paginiDevelopment of The MaxillaShinta Syafe'iÎncă nu există evaluări
- Smell & TasteDocument7 paginiSmell & TastePatterson MachariaÎncă nu există evaluări
- Moisture ControlDocument43 paginiMoisture ControlPatterson Macharia100% (1)
- Higher Functions of The Nervous SystemDocument8 paginiHigher Functions of The Nervous SystemPatterson MachariaÎncă nu există evaluări
- Alert Behavior, Sleep and The Electrical Activity of The BrainDocument6 paginiAlert Behavior, Sleep and The Electrical Activity of The BrainPatterson MachariaÎncă nu există evaluări
- Hearing and EquilibriumDocument11 paginiHearing and EquilibriumPatterson MachariaÎncă nu există evaluări
- ReflexesDocument6 paginiReflexesPatterson MachariaÎncă nu există evaluări
- Cutaneous, Deep and Visceral SensationDocument12 paginiCutaneous, Deep and Visceral SensationPatterson MachariaÎncă nu există evaluări
- Neural Basis of Instinctual BehaviorDocument8 paginiNeural Basis of Instinctual BehaviorPatterson MachariaÎncă nu există evaluări
- CNS PharmacologyDocument26 paginiCNS PharmacologyPatterson MachariaÎncă nu există evaluări
- Adrenergic Receptor AntagonistsDocument9 paginiAdrenergic Receptor AntagonistsPatterson MachariaÎncă nu există evaluări
- Central Regulation of Visceral FunctionDocument6 paginiCentral Regulation of Visceral FunctionPatterson MachariaÎncă nu există evaluări
- Does Sastra Prescribes Animal Sacrifices For The Sake of ReligionDocument15 paginiDoes Sastra Prescribes Animal Sacrifices For The Sake of Religionkeshav3kashmiriÎncă nu există evaluări
- Antiprotozoal Agents 6Document29 paginiAntiprotozoal Agents 6EinsteenÎncă nu există evaluări
- A Detailed Lesson Plan in Partial Fulfillment of The Course Requirements in (PED8)Document11 paginiA Detailed Lesson Plan in Partial Fulfillment of The Course Requirements in (PED8)Angel Clarence BasilioÎncă nu există evaluări
- Module Possessive Adj. Lesson 3Document29 paginiModule Possessive Adj. Lesson 3Lance TuvillaÎncă nu există evaluări
- July Aug MenuDocument1 paginăJuly Aug Menusales6916Încă nu există evaluări
- Nursery RhymesDocument10 paginiNursery RhymesDegee O. GonzalesÎncă nu există evaluări
- Removable Partial DentureDocument4 paginiRemovable Partial DentureRodriguez VanesaÎncă nu există evaluări
- American Rabbit Breeders Association Guide (ARBADocument11 paginiAmerican Rabbit Breeders Association Guide (ARBANaniÎncă nu există evaluări
- Plitvice Lakes, CroatiaDocument7 paginiPlitvice Lakes, CroatiaMirkoÎncă nu există evaluări
- Catclass NotesDocument3 paginiCatclass NotesIssac CaldwellÎncă nu există evaluări
- Fluency Passages P1Document99 paginiFluency Passages P1princerafa100% (5)
- Animal Science HandoutDocument5 paginiAnimal Science Handoutromnickatchecoso14Încă nu există evaluări
- Normal Adult Laboratory ValuesDocument7 paginiNormal Adult Laboratory Valuessteinway007Încă nu există evaluări
- Organ DonationDocument3 paginiOrgan DonationGrant DonovanÎncă nu există evaluări
- Vaults of Vaarn #1 (v3) (Booklet) (2021-01-11)Document26 paginiVaults of Vaarn #1 (v3) (Booklet) (2021-01-11)Alex TrifanÎncă nu există evaluări
- ECG BasicsDocument15 paginiECG Basicsrohitmeena2889Încă nu există evaluări
- Histology AssignmentDocument4 paginiHistology AssignmentNico LokoÎncă nu există evaluări
- Y1 Module Pet ShowDocument13 paginiY1 Module Pet ShowJayne Frances EngelbertÎncă nu există evaluări
- The Little SquirrelDocument10 paginiThe Little SquirrelNgọc Ngọc100% (10)
- Ultimate Arcane SpellbookDocument32 paginiUltimate Arcane SpellbookSigraine75% (4)
- AnatomyDocument4 paginiAnatomySureen PaduaÎncă nu există evaluări
- Spiral FlapDocument20 paginiSpiral FlapDiolanda De AbreuÎncă nu există evaluări
- The Sign of The BeaverDocument13 paginiThe Sign of The Beaverjuan marcos100% (1)
- Acute Respiratory InfectionDocument32 paginiAcute Respiratory InfectionRinnaAyuNovitaSary100% (1)
- Science Notes Sem1 PDFDocument31 paginiScience Notes Sem1 PDFWolfie Saraswathi Santhosham100% (1)
- 2007 IGCSE Biology Written Paper Question PaperDocument24 pagini2007 IGCSE Biology Written Paper Question Paperseyka4Încă nu există evaluări
- Shark Finning Report WildaidDocument16 paginiShark Finning Report WildaidGlobal Wildlife Warriors100% (1)
- Eduardo Serrão - Potential Business For Chickens and Pigs and Their Products in Timor-LesteDocument6 paginiEduardo Serrão - Potential Business For Chickens and Pigs and Their Products in Timor-LestePapers and Powerpoints from UNTL-VU Joint Conferenes in DiliÎncă nu există evaluări
- A - Specialist Catalogue Fall 2019-Web PDFDocument29 paginiA - Specialist Catalogue Fall 2019-Web PDFFelipe YacumalÎncă nu există evaluări
- The Age of Magical Overthinking: Notes on Modern IrrationalityDe la EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityEvaluare: 4 din 5 stele4/5 (13)
- Techniques Exercises And Tricks For Memory ImprovementDe la EverandTechniques Exercises And Tricks For Memory ImprovementEvaluare: 4.5 din 5 stele4.5/5 (40)
- The Ultimate Guide To Memory Improvement TechniquesDe la EverandThe Ultimate Guide To Memory Improvement TechniquesEvaluare: 5 din 5 stele5/5 (34)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeDe la EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeÎncă nu există evaluări
- The Happiness Trap: How to Stop Struggling and Start LivingDe la EverandThe Happiness Trap: How to Stop Struggling and Start LivingEvaluare: 4 din 5 stele4/5 (1)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedDe la EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedEvaluare: 5 din 5 stele5/5 (78)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionDe la EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionEvaluare: 4 din 5 stele4/5 (402)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDe la EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityEvaluare: 3.5 din 5 stele3.5/5 (2)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsDe la EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsEvaluare: 3.5 din 5 stele3.5/5 (3)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsDe la EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsEvaluare: 4.5 din 5 stele4.5/5 (169)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsDe la EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsEvaluare: 5 din 5 stele5/5 (1)
- The Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsDe la EverandThe Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsÎncă nu există evaluări
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaDe la EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaEvaluare: 4.5 din 5 stele4.5/5 (266)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessDe la EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessEvaluare: 4.5 din 5 stele4.5/5 (327)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisDe la EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisEvaluare: 4 din 5 stele4/5 (1)
- The Tennis Partner: A Doctor's Story of Friendship and LossDe la EverandThe Tennis Partner: A Doctor's Story of Friendship and LossEvaluare: 4.5 din 5 stele4.5/5 (4)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.De la EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Evaluare: 4.5 din 5 stele4.5/5 (110)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryDe la EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryEvaluare: 4 din 5 stele4/5 (44)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeDe la EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeEvaluare: 4.5 din 5 stele4.5/5 (253)
- The Stress-Proof Brain: Master Your Emotional Response to Stress Using Mindfulness and NeuroplasticityDe la EverandThe Stress-Proof Brain: Master Your Emotional Response to Stress Using Mindfulness and NeuroplasticityEvaluare: 4.5 din 5 stele4.5/5 (109)
- Secure Love: Create a Relationship That Lasts a LifetimeDe la EverandSecure Love: Create a Relationship That Lasts a LifetimeEvaluare: 5 din 5 stele5/5 (17)
- Summary: It Didn't Start with You: How Inherited Family Trauma Shapes Who We Are and How to End the Cycle By Mark Wolynn: Key Takeaways, Summary & AnalysisDe la EverandSummary: It Didn't Start with You: How Inherited Family Trauma Shapes Who We Are and How to End the Cycle By Mark Wolynn: Key Takeaways, Summary & AnalysisEvaluare: 5 din 5 stele5/5 (3)
- Summary: Hidden Potential: The Science of Achieving Greater Things By Adam Grant: Key Takeaways, Summary and AnalysisDe la EverandSummary: Hidden Potential: The Science of Achieving Greater Things By Adam Grant: Key Takeaways, Summary and AnalysisEvaluare: 4.5 din 5 stele4.5/5 (15)
- Troubled: A Memoir of Foster Care, Family, and Social ClassDe la EverandTroubled: A Memoir of Foster Care, Family, and Social ClassEvaluare: 4.5 din 5 stele4.5/5 (23)
- Hearts of Darkness: Serial Killers, The Behavioral Science Unit, and My Life as a Woman in the FBIDe la EverandHearts of Darkness: Serial Killers, The Behavioral Science Unit, and My Life as a Woman in the FBIEvaluare: 4 din 5 stele4/5 (19)
- Summary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisDe la EverandSummary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisEvaluare: 5 din 5 stele5/5 (8)
- Daniel Kahneman's "Thinking Fast and Slow": A Macat AnalysisDe la EverandDaniel Kahneman's "Thinking Fast and Slow": A Macat AnalysisEvaluare: 3.5 din 5 stele3.5/5 (130)