S-ar putea să vă placă și
- Alytes, 16 (1-2), 61-67 - CrossodactylusDocument7 paginiAlytes, 16 (1-2), 61-67 - CrossodactylusSebas CirignoliÎncă nu există evaluări
- Systematic Status of a South American Frog, Allophryne ruthveni GaigeDe la EverandSystematic Status of a South American Frog, Allophryne ruthveni GaigeÎncă nu există evaluări
- 10 Nunes e Pombal S JuncaeDocument7 pagini10 Nunes e Pombal S JuncaeJoseane de Souza CardosoÎncă nu există evaluări
- 2007 Bryophryne Bustamantei Chaparro EtalDocument8 pagini2007 Bryophryne Bustamantei Chaparro EtalJuanca Chaparro AuzaÎncă nu există evaluări
- A New Species of From The Juan Ferna Ndez Islands: Eupithecia (Lepidoptera: Geometridae)Document7 paginiA New Species of From The Juan Ferna Ndez Islands: Eupithecia (Lepidoptera: Geometridae)Héctor Ibarra VidalÎncă nu există evaluări
- Torres-Carvajal2005-A New Species of Stenocercus (Squamata Iguanidae) From Central-Western Brazil With A Key To BraziDocument10 paginiTorres-Carvajal2005-A New Species of Stenocercus (Squamata Iguanidae) From Central-Western Brazil With A Key To BraziCristhian Raúl Rossi La TorreÎncă nu există evaluări
- Guerrero 2007Document23 paginiGuerrero 2007tqphapÎncă nu există evaluări
- Haverkost and Gardner 2008 ADocument8 paginiHaverkost and Gardner 2008 At_haverkostÎncă nu există evaluări
- LobezaDocument8 paginiLobezaEmerson PawoskiÎncă nu există evaluări
- Helminth Parasites of Two Sympatric Lizards, From An Atlantic Rainforest Area of Southeastern BrazilDocument4 paginiHelminth Parasites of Two Sympatric Lizards, From An Atlantic Rainforest Area of Southeastern BrazilWillison Do CarmoÎncă nu există evaluări
- Asimos Staphylinidae SudamericaDocument11 paginiAsimos Staphylinidae SudamericaWalter Guillermo Cosio LoaizaÎncă nu există evaluări
- A New Brachypterous Mesepipsocus (Psocodea: 'Psocoptera': Epipsocidae) From BoliviaDocument4 paginiA New Brachypterous Mesepipsocus (Psocodea: 'Psocoptera': Epipsocidae) From Boliviaonurb90Încă nu există evaluări
- Artigo Ninfa AnacroneuriaDocument16 paginiArtigo Ninfa AnacroneuriaTainah OliveiraÎncă nu există evaluări
- New Species of Melanophryniscus (Anura: Bufonidae) From Rio Grande Do Sul, Southern BrazilDocument6 paginiNew Species of Melanophryniscus (Anura: Bufonidae) From Rio Grande Do Sul, Southern BrazilAna AriasÎncă nu există evaluări
- Linkem Et Al.2008 PDFDocument11 paginiLinkem Et Al.2008 PDFheru2910Încă nu există evaluări
- Burs Ey Kinsella 2003Document5 paginiBurs Ey Kinsella 2003Mohamad JakariaÎncă nu există evaluări
- Corallus Blombergi Notes From Ecuador Valencia Et Al. 2008Document4 paginiCorallus Blombergi Notes From Ecuador Valencia Et Al. 2008Samir ValenciaÎncă nu există evaluări
- The Tadpole of Bokermannohyla CapraDocument3 paginiThe Tadpole of Bokermannohyla CapraAiran ProtázioÎncă nu există evaluări
- Description of The Tadpole of Megaelosia Goeldii (Leptodactylidae, Hylodinae) With Natural HistorDocument6 paginiDescription of The Tadpole of Megaelosia Goeldii (Leptodactylidae, Hylodinae) With Natural HistorJoseane de Souza CardosoÎncă nu există evaluări
- 2017 Marciano-Jr Et Al. Phyllodytes MegatimpanumDocument8 pagini2017 Marciano-Jr Et Al. Phyllodytes MegatimpanumamandasantiagoÎncă nu există evaluări
- Martins 2019Document11 paginiMartins 2019Luciana MartinsÎncă nu există evaluări
- Bicentenariella Sinuosa Serranid Fishes New Species 2021 Parecida A Monogeneo de Cichlasoma IstlanumDocument11 paginiBicentenariella Sinuosa Serranid Fishes New Species 2021 Parecida A Monogeneo de Cichlasoma IstlanumBelem FloresÎncă nu există evaluări
- Bonvicino 2002Document11 paginiBonvicino 2002Maria EduardaÎncă nu există evaluări
- A New Snake of the Genus Geophis From Chihuahua, MexicoDe la EverandA New Snake of the Genus Geophis From Chihuahua, MexicoÎncă nu există evaluări
- MendozamirisDocument6 paginiMendozamirisLan YenchiuÎncă nu există evaluări
- 2013 Lourenço-De-Moraes Et Al. Girino e Canto Aparasphenodon ArapapaDocument5 pagini2013 Lourenço-De-Moraes Et Al. Girino e Canto Aparasphenodon ArapapaamandasantiagoÎncă nu există evaluări
- A Taxonomic Study of the Middle American Snake, Pituophis deppeiDe la EverandA Taxonomic Study of the Middle American Snake, Pituophis deppeiÎncă nu există evaluări
- Artigo Parasito GambaDocument7 paginiArtigo Parasito GambaFernanda RezendeÎncă nu există evaluări
- Dos Nuevas Especies de PhyllodactylusDocument12 paginiDos Nuevas Especies de PhyllodactylusLuis F. GarcíaÎncă nu există evaluări
- Ma 2006 J Euk Micro Epistylis Onblue CrabDocument11 paginiMa 2006 J Euk Micro Epistylis Onblue CrabRIZKI NANDAÎncă nu există evaluări
- The Sphaerodactylus (Sauria Gekkonidae) of South America PDFDocument32 paginiThe Sphaerodactylus (Sauria Gekkonidae) of South America PDFanon_243369186Încă nu există evaluări
- Avicularia Rickwesti Sp. Nov., A Remarkable New Species of AviculariaDocument5 paginiAvicularia Rickwesti Sp. Nov., A Remarkable New Species of AviculariazzzaÎncă nu există evaluări
- A New Species of Pseudapocyrtus H, 1912 (Coleoptera: Curculionidae) From Southern Luzon, The PhilippinesDocument6 paginiA New Species of Pseudapocyrtus H, 1912 (Coleoptera: Curculionidae) From Southern Luzon, The PhilippinesHajime HikariÎncă nu există evaluări
- Revision of Philodryas Mattogrossensis With The Revalidation of P. Erlandi (Reptilia: Squamata: Dipsadidae)Document15 paginiRevision of Philodryas Mattogrossensis With The Revalidation of P. Erlandi (Reptilia: Squamata: Dipsadidae)Daniela CerqueiraÎncă nu există evaluări
- Morphology, Reproductive Biology and Diet of Colubridae) in Brazilian AmazonDocument7 paginiMorphology, Reproductive Biology and Diet of Colubridae) in Brazilian AmazonEdiegoBatistaÎncă nu există evaluări
- The Genera of Phyllomedusine Frogs (Anura Hylidae)De la EverandThe Genera of Phyllomedusine Frogs (Anura Hylidae)Încă nu există evaluări
- 1977 Lynch & Hoogmoed - Two New Eleutherodactylus (Chiastonotus, Zeuctotylus) From NE South AmericaDocument17 pagini1977 Lynch & Hoogmoed - Two New Eleutherodactylus (Chiastonotus, Zeuctotylus) From NE South AmericaMarinus HoogmoedÎncă nu există evaluări
- Records of Harvest Mice, Reithrodontomys, from Central America, with Description of a New Subspecies from NicaraguaDe la EverandRecords of Harvest Mice, Reithrodontomys, from Central America, with Description of a New Subspecies from NicaraguaÎncă nu există evaluări
- Reptilia, Squamata, Amphisbaenidae, Leposternon SPP.: Distribution Extension, New State Record, and Geographic Distribution MapDocument4 paginiReptilia, Squamata, Amphisbaenidae, Leposternon SPP.: Distribution Extension, New State Record, and Geographic Distribution MapkellymafraÎncă nu există evaluări
- Rangel Santos 2009 JParasitol 95 5 1233-1236Document4 paginiRangel Santos 2009 JParasitol 95 5 1233-1236totilas2112Încă nu există evaluări
- Southernmost Records of Hyalinobatrachium Fleischmanni (Anura: Centrolenidae)Document4 paginiSouthernmost Records of Hyalinobatrachium Fleischmanni (Anura: Centrolenidae)Juan Carlos SánchezÎncă nu există evaluări
- Torres Carvajal 2005bDocument7 paginiTorres Carvajal 2005bjoel ayala navarroÎncă nu există evaluări
- Culebras Etal 2023Document4 paginiCulebras Etal 2023s6amplewÎncă nu există evaluări
- ZK Article 49434 en 1Document12 paginiZK Article 49434 en 1Walter Guillermo Cosio LoaizaÎncă nu există evaluări
- A Taxonomic Study of the Middle American Snake, Pituophis deppeiDe la EverandA Taxonomic Study of the Middle American Snake, Pituophis deppeiÎncă nu există evaluări
- Cultivo Harknessia de EucaliptoDocument18 paginiCultivo Harknessia de EucaliptoRosales Rosales JesúsÎncă nu există evaluări
- Cocceupodidae, A New Family of Eupodoid Mites, With Description of A New Genus and Two New Species From Poland. Part I. (Acari: Prostigmata: Eupodoidea)Document22 paginiCocceupodidae, A New Family of Eupodoid Mites, With Description of A New Genus and Two New Species From Poland. Part I. (Acari: Prostigmata: Eupodoidea)Je suis un MicaschisteÎncă nu există evaluări
- First Record of Aplatophis Chauliodus (Actinopterygii: Ophichthidae) For The Northeastern Coast of Brazil, With Expansion of Its Geographic RangeDocument4 paginiFirst Record of Aplatophis Chauliodus (Actinopterygii: Ophichthidae) For The Northeastern Coast of Brazil, With Expansion of Its Geographic RangeBuia baianoÎncă nu există evaluări
- Osteocephalus 3 SP Nov 2012Document52 paginiOsteocephalus 3 SP Nov 2012Aleksandra PanyutinaÎncă nu există evaluări
- Adelophryne AdiastolaDocument5 paginiAdelophryne Adiastolajonho2006Încă nu există evaluări
- 213 803 1 SMDocument16 pagini213 803 1 SMmarcio vieiraÎncă nu există evaluări
- Condez Etal2014 Bcrispus PDFDocument12 paginiCondez Etal2014 Bcrispus PDFvanessa e bruno becaciciÎncă nu există evaluări
- A New Treefrog of The Genus Rhacophorus (Anura: Rhacophoridae) From Brunei Darussalam (Borneo)Document12 paginiA New Treefrog of The Genus Rhacophorus (Anura: Rhacophoridae) From Brunei Darussalam (Borneo)Budi AfriyansyahÎncă nu există evaluări
- Kyphosis and other Variations in Soft-shelled TurtlesDe la EverandKyphosis and other Variations in Soft-shelled TurtlesÎncă nu există evaluări
- Deforeletal 2021melanophryniscusDocument27 paginiDeforeletal 2021melanophryniscusjose.1711717678Încă nu există evaluări
- DesjardinPerry Mycosphere7 2016Document33 paginiDesjardinPerry Mycosphere7 2016Fernando FabriniÎncă nu există evaluări
- Protocylindrocorpus BrasiliensisDocument6 paginiProtocylindrocorpus BrasiliensisAzhari RizalÎncă nu există evaluări
- Description of A New Species of Australian Cerdistus (Asilidae: Asilinae)Document14 paginiDescription of A New Species of Australian Cerdistus (Asilidae: Asilinae)TommasoÎncă nu există evaluări
- Torquatus (Wied-Neuwied, 1820) (Sauria: Tropiduridae)Document3 paginiTorquatus (Wied-Neuwied, 1820) (Sauria: Tropiduridae)Alexander RochaÎncă nu există evaluări
- First Record of Attempted Piscivory by Bothrops Asper (Garman, 1883) (Squamata, Viperidae) On A Swamp Eel, Genus SynbranchusDocument3 paginiFirst Record of Attempted Piscivory by Bothrops Asper (Garman, 1883) (Squamata, Viperidae) On A Swamp Eel, Genus SynbranchusJCAÎncă nu există evaluări
- Boessenecker Et Al-2015-Zoological Journal of The Linnean SocietyDocument54 paginiBoessenecker Et Al-2015-Zoological Journal of The Linnean SocietyJCAÎncă nu există evaluări
- 6821 21083 1 PBDocument2 pagini6821 21083 1 PBJCAÎncă nu există evaluări
- Vicariance and Endemism in A Neotropical Savanna Hotspot - Distribution Patterns of Cerrado Squamate ReptilesDocument16 paginiVicariance and Endemism in A Neotropical Savanna Hotspot - Distribution Patterns of Cerrado Squamate ReptilesJCAÎncă nu există evaluări
- GekoDocument22 paginiGekoVíctor MundoÎncă nu există evaluări
- Harmful Algal Blooms and Eutrophication Nutrient Sources, Composition, and ConsequencesDocument23 paginiHarmful Algal Blooms and Eutrophication Nutrient Sources, Composition, and ConsequencesJCAÎncă nu există evaluări
- Anesthesia CPT and Icd 9 CodesDocument36 paginiAnesthesia CPT and Icd 9 CodesSundarajan ManiÎncă nu există evaluări
- Untitled 2 PDFDocument10 paginiUntitled 2 PDFspike9444Încă nu există evaluări
- Part One 2008-2018 by Saud AlmaslmaniDocument108 paginiPart One 2008-2018 by Saud AlmaslmaniAlHasa100% (5)
- Anaphy Lab Oral Recitation 1-7Document10 paginiAnaphy Lab Oral Recitation 1-7Niña C.Încă nu există evaluări
- Kinesiology Exam ReviewDocument13 paginiKinesiology Exam Reviewazn_taiwen8100% (1)
- A Simplified Approach To Nasal Tip Shaping: The Five-Suture TechniqueDocument5 paginiA Simplified Approach To Nasal Tip Shaping: The Five-Suture TechniqueroyvillafrancaÎncă nu există evaluări
- Medical Terminology PresentationDocument68 paginiMedical Terminology Presentationasem sawalmehÎncă nu există evaluări
- Border MoldingDocument39 paginiBorder MoldingvsdeepsÎncă nu există evaluări
- Muscle Length TestDocument7 paginiMuscle Length Testsigue_brigitteangeli_lanotÎncă nu există evaluări
- Injury: Orlin Filipov, Boyko GueorguievDocument9 paginiInjury: Orlin Filipov, Boyko GueorguievHein Aung ZawÎncă nu există evaluări
- People v. Pagador PDFDocument10 paginiPeople v. Pagador PDFAAMCÎncă nu există evaluări
- Deformaciones FlexuralesDocument11 paginiDeformaciones FlexuralesCamila BragagniniÎncă nu există evaluări
- Introodu Ction: Terminology PDFDocument3 paginiIntroodu Ction: Terminology PDFAmirah AnfÎncă nu există evaluări
- Venous DiseaseDocument45 paginiVenous DiseaseNinch Nagac100% (1)
- Anatomy of The Breast: Alison J. DarlingtonDocument8 paginiAnatomy of The Breast: Alison J. DarlingtonValir HusleÎncă nu există evaluări
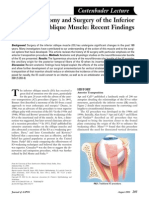
- Inferior Oblique MuscleDocument6 paginiInferior Oblique MuscleYunita ManurungÎncă nu există evaluări
- Manufacturing & Fitting The Jaipur Below-the-Knee ProsthesisDocument69 paginiManufacturing & Fitting The Jaipur Below-the-Knee ProsthesisoxideheraÎncă nu există evaluări
- Leseprobe Akupunktur Atlas GB HfullmannDocument13 paginiLeseprobe Akupunktur Atlas GB Hfullmannalvaedison00Încă nu există evaluări
- Charting Examples For Physical AssessmentDocument16 paginiCharting Examples For Physical Assessmentim3in1Încă nu există evaluări
- Dpem Ort 1212 0405 - 9074 50 001 - LRDocument20 paginiDpem Ort 1212 0405 - 9074 50 001 - LRyvesÎncă nu există evaluări
- Principles of Radiographic Interpretation: Submitted By:-Saikat Kumar Kundu MVSC 2 Year ID-54187Document104 paginiPrinciples of Radiographic Interpretation: Submitted By:-Saikat Kumar Kundu MVSC 2 Year ID-54187Praney SlathiaÎncă nu există evaluări
- WKoTaXFrZtNGz2xzB - Seeleys Ch1Document43 paginiWKoTaXFrZtNGz2xzB - Seeleys Ch1Björn Þór SigurbjörnssonÎncă nu există evaluări
- Appendix 3: SPCM Scoring Form: Sunny Hill Health Centre For Children 3644 Slocan Street, Vancouver, B.C. V5M 3E8Document6 paginiAppendix 3: SPCM Scoring Form: Sunny Hill Health Centre For Children 3644 Slocan Street, Vancouver, B.C. V5M 3E8Laura Vanessa Caballero ArizaÎncă nu există evaluări
- A Dissection Course On Endoscopic Endonasal Sinus Surgery PDFDocument36 paginiA Dissection Course On Endoscopic Endonasal Sinus Surgery PDFLupita Marquez100% (1)
- Femoral NerveDocument11 paginiFemoral NerveJhon QuirogaÎncă nu există evaluări
- Wood, E. J. F. 1963. Dinoflagellates in The Australian RegionDocument55 paginiWood, E. J. F. 1963. Dinoflagellates in The Australian RegionAlejandro EstewesonÎncă nu există evaluări
- Description Dermatological StatusDocument6 paginiDescription Dermatological StatusNazmun LailahÎncă nu există evaluări
- Laquan McDonald Police Reports, Part 3Document60 paginiLaquan McDonald Police Reports, Part 3jroneillÎncă nu există evaluări
- Posterior Palatal SealDocument37 paginiPosterior Palatal Sealreem eltyebÎncă nu există evaluări
- LABORATORY MANUAL in GENERAL ANATOMY 1 PART 1Document29 paginiLABORATORY MANUAL in GENERAL ANATOMY 1 PART 1Ian Mar ValenzuelaÎncă nu există evaluări