S-ar putea să vă placă și
- Asegura tu maternidad: Todo lo que necesitas saber sobre el congelamiento de óvulosDe la EverandAsegura tu maternidad: Todo lo que necesitas saber sobre el congelamiento de óvulosÎncă nu există evaluări
- Trabajo BiotecnologiaDocument24 paginiTrabajo BiotecnologiasoruizÎncă nu există evaluări
- Lopez - Desarrollo de Embriones de Conejo Sobre Matriz Oviductal y Medio Condicionado Por Celulas...Document61 paginiLopez - Desarrollo de Embriones de Conejo Sobre Matriz Oviductal y Medio Condicionado Por Celulas...sam SalinasÎncă nu există evaluări
- Tesis Trucha PDFDocument46 paginiTesis Trucha PDFmeyerhuaranga100% (3)
- Practica Ovocitos PDFDocument10 paginiPractica Ovocitos PDFAdan MenesesÎncă nu există evaluări
- PRACTICA Nº6 Recuperación de OvulosDocument8 paginiPRACTICA Nº6 Recuperación de Ovulosmili-taÎncă nu există evaluări
- Tesis para ImprimirDocument139 paginiTesis para Imprimirisabel velezÎncă nu există evaluări
- Mi TesisDocument96 paginiMi TesisAdrian Betanzos RodriguezÎncă nu există evaluări
- Para El ProyectoDocument150 paginiPara El ProyectoMISHEL ROSARIO ROMERO MARAVIÎncă nu există evaluări
- Biocells Células Madre 2014Document12 paginiBiocells Células Madre 2014William GonzálezÎncă nu există evaluări
- Evaluación Del Rendimiento Productivo de LasDocument116 paginiEvaluación Del Rendimiento Productivo de LasCarlos Beltran ChaconÎncă nu există evaluări
- Bejarano-Álvarez,+M +2007 +Tesis+de+Maestría TiburonesDocument93 paginiBejarano-Álvarez,+M +2007 +Tesis+de+Maestría TiburonesSalvador PeraltaÎncă nu există evaluări
- Articulos Cientificos de AlpacasDocument3 paginiArticulos Cientificos de AlpacasAndyMHuisaÎncă nu există evaluări
- Evaluacion de Protoolo de Transferencia de EmbrionesDocument25 paginiEvaluacion de Protoolo de Transferencia de EmbrionesDiego AscuntarÎncă nu există evaluări
- Desarrollo Embrionario Temprano en El HamsterDocument11 paginiDesarrollo Embrionario Temprano en El HamsterStephany Lochi AguilarÎncă nu există evaluări
- Embarazo Multiple - ObstetriciaDocument20 paginiEmbarazo Multiple - ObstetriciaMario MachadoÎncă nu există evaluări
- 34 Alarico Zeballos DA FCAG Medicina Veterinaria Zootecnia 2012 PDFDocument134 pagini34 Alarico Zeballos DA FCAG Medicina Veterinaria Zootecnia 2012 PDFjhon elíÎncă nu există evaluări
- Transferencia de Embriones Erick Ramos ZeballosDocument14 paginiTransferencia de Embriones Erick Ramos ZeballosErick RamosÎncă nu există evaluări
- Pubertad y Circunferencia Escrotal en Toros Holstein X CebuDocument4 paginiPubertad y Circunferencia Escrotal en Toros Holstein X CebuMaria Fernanda IriarteÎncă nu există evaluări
- Biologia Practica 1Document11 paginiBiologia Practica 1illojuanjuniorÎncă nu există evaluări
- Texto Cultivo in VitroDocument43 paginiTexto Cultivo in VitroYonatan MamaniÎncă nu există evaluări
- Evaluación Citogenética Del CCCDocument73 paginiEvaluación Citogenética Del CCCAlejandroNietoÎncă nu există evaluări
- Tolentino Castro C MC Ganaderia 2014Document61 paginiTolentino Castro C MC Ganaderia 2014Erick GuzmanÎncă nu există evaluări
- Produccion in Vitro en Embriones BovinosDocument9 paginiProduccion in Vitro en Embriones BovinosprincipelicÎncă nu există evaluări
- Evaluacion de La Crioconservacion de Embriones Bovinos (Document37 paginiEvaluacion de La Crioconservacion de Embriones Bovinos (Danilo Lobo CalleÎncă nu există evaluări
- Somatotropina Bovina Recombinante yDocument63 paginiSomatotropina Bovina Recombinante ysalvador alonsoÎncă nu există evaluări
- Congelacion y Sexado Semen BovinoDocument6 paginiCongelacion y Sexado Semen BovinoFelipe ArevaloÎncă nu există evaluări
- Curva de Crecimiento B. SubtilisDocument3 paginiCurva de Crecimiento B. SubtilisGuadalupe FloresÎncă nu există evaluări
- Practica 6 Recolección de Ovocitos E.E PDFDocument3 paginiPractica 6 Recolección de Ovocitos E.E PDFXDianaCrznXCharajaÎncă nu există evaluări
- Tesis Juan Carlos Flores RiosDocument74 paginiTesis Juan Carlos Flores RiosJ'CarlosFloresÎncă nu există evaluări
- Monografía MetodosDocument17 paginiMonografía MetodosRoshiory Teresa Huamán ZamalloaÎncă nu există evaluări
- Semana 06 - Produccion in Vitro de Embriones BovinosDocument44 paginiSemana 06 - Produccion in Vitro de Embriones BovinosJorge Janampa CamposÎncă nu există evaluări
- MONOGRAFIA LEONARDO - Sincornizacion de Partos en CerdasDocument32 paginiMONOGRAFIA LEONARDO - Sincornizacion de Partos en CerdasAnahí RetamozoÎncă nu există evaluări
- Desarrollo Embrionario de Danio Rerio Oficial Del JuevesDocument15 paginiDesarrollo Embrionario de Danio Rerio Oficial Del JuevesSalomon ZegarraÎncă nu există evaluări
- Desarrollo Embrionario de Chicoreus BrevifronsDocument99 paginiDesarrollo Embrionario de Chicoreus BrevifronsEnioÎncă nu există evaluări
- Bolilla 5Document25 paginiBolilla 5Juan LucasÎncă nu există evaluări
- Tesis Lic Zoot Hersson Giovanny Icú Cutzal PDFDocument39 paginiTesis Lic Zoot Hersson Giovanny Icú Cutzal PDFVictorÎncă nu există evaluări
- Gyna Acosta VillotaDocument116 paginiGyna Acosta VillotaRONALD HERNANDEZÎncă nu există evaluări
- BIOTECNOLOGIADocument14 paginiBIOTECNOLOGIADanilo Lobo CalleÎncă nu există evaluări
- Upse Tia 2021 0090Document57 paginiUpse Tia 2021 0090Elvis Aviles MendozaÎncă nu există evaluări
- Producion in - Vitro de Emb TitiDocument33 paginiProducion in - Vitro de Emb TitiEdwin A. Aguilar M.Încă nu există evaluări
- Efecto Cicatrizante Del Ungüento A Base deDocument143 paginiEfecto Cicatrizante Del Ungüento A Base dePaul Mendoza SalasÎncă nu există evaluări
- Determination of Technological Parameters For The Preparation ofDocument305 paginiDetermination of Technological Parameters For The Preparation ofDenis Huanca De La CruzÎncă nu există evaluări
- Monografia ClonacionDocument29 paginiMonografia ClonacionYessi CYÎncă nu există evaluări
- 1609 9117 Rivep 34 02 E25101Document17 pagini1609 9117 Rivep 34 02 E25101rbarretorÎncă nu există evaluări
- Modelo de La Presentacion Del Informe de PracticaDocument3 paginiModelo de La Presentacion Del Informe de PracticaWilber Julca SosaÎncă nu există evaluări
- Manual para Hacer CorredoresDocument220 paginiManual para Hacer Corredoresmarco pinto100% (1)
- Fisiología Reproductiva Del Pez Betta Splendens en Condiciones de Laboratorio, Piedemonte Andino Amazónico (Colombia)Document9 paginiFisiología Reproductiva Del Pez Betta Splendens en Condiciones de Laboratorio, Piedemonte Andino Amazónico (Colombia)EliamAlejandraArdilaMorenoÎncă nu există evaluări
- Ancova CamaronDocument178 paginiAncova CamaronMaritza RodriguezÎncă nu există evaluări
- Calidad Del Semen en Trucha Arco Iris (Onchorhynchus Mykiss) Lote Michoacano, A Finales de Su Periodo ReproductivoDocument10 paginiCalidad Del Semen en Trucha Arco Iris (Onchorhynchus Mykiss) Lote Michoacano, A Finales de Su Periodo ReproductivoDarGinn Salamanca CÎncă nu există evaluări
- Tesis para Sustentación Pública-Sebastian VinuezaDocument68 paginiTesis para Sustentación Pública-Sebastian VinuezaSebastian Jaén Vinueza SoteloÎncă nu există evaluări
- Astudillo Carolina 1317dDocument250 paginiAstudillo Carolina 1317dSEBASTIÁN IGNACIO OÑATEÎncă nu există evaluări
- Diametro de OvocitosDocument3 paginiDiametro de OvocitosSergio FerÎncă nu există evaluări
- Soto Valenzuela Javier OswaldoDocument166 paginiSoto Valenzuela Javier OswaldoNORMA ELIANA BARBAGGELATA CHALCOÎncă nu există evaluări
- P2. Curva de Crecimiento BacterianoDocument7 paginiP2. Curva de Crecimiento BacterianoJhohanÎncă nu există evaluări
- Alvarez Ye - DesbloqueadoDocument169 paginiAlvarez Ye - DesbloqueadoJesús Alberto Carrasco BocanegraÎncă nu există evaluări
- Esquema para El Informe de PrácticaDocument4 paginiEsquema para El Informe de PrácticaARIANA ESTRELLA ZUÑIGA MANRIQUEÎncă nu există evaluări
- Ganancia de Peso (Morera)Document78 paginiGanancia de Peso (Morera)David PalomaresÎncă nu există evaluări
- Comportamiento productivo y reproductivo en caprinos en trópico secoDe la EverandComportamiento productivo y reproductivo en caprinos en trópico secoEvaluare: 1 din 5 stele1/5 (1)
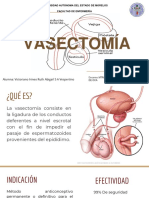
- VASECTOMÍADocument9 paginiVASECTOMÍARuth Abigail Victoriano IrineoÎncă nu există evaluări
- Docencia, Rol e IdentidadDocument3 paginiDocencia, Rol e IdentidadJuan Mata PimentelÎncă nu există evaluări
- Plantilla Presentación Gratis GalvestonDocument28 paginiPlantilla Presentación Gratis GalvestonDiana Paola Garcia FigueroaÎncă nu există evaluări
- Sesión 3 Diferencias y Semejanzas Entre La Contabilidad Gerencial y La Contabilidad FinancieraDocument31 paginiSesión 3 Diferencias y Semejanzas Entre La Contabilidad Gerencial y La Contabilidad FinancieraJEHYNER JESUS HUARACA GOITIAÎncă nu există evaluări
- 16b. Petrarca - Subida Al Monte VentosoDocument4 pagini16b. Petrarca - Subida Al Monte VentosoMauricio Thunders Contreras50% (2)
- Tesis FinalDocument134 paginiTesis FinalbarcesalaÎncă nu există evaluări
- Caso Práctico N2 - Gestión de La CalidadDocument3 paginiCaso Práctico N2 - Gestión de La CalidadAngela Rocha HernandezÎncă nu există evaluări
- Trabajo Final Ingri 2Document28 paginiTrabajo Final Ingri 2SailethÎncă nu există evaluări
- Calculo ChavetaDocument5 paginiCalculo ChavetaJoscarin BalandÎncă nu există evaluări
- Reglamento Modalidades para Educación Especial FinalDocument11 paginiReglamento Modalidades para Educación Especial FinalPABKAÎncă nu există evaluări
- Resumen Certamen 1Document4 paginiResumen Certamen 1nacho popÎncă nu există evaluări
- Hierba Del ClavoDocument2 paginiHierba Del ClavoGentil GoyimwitzÎncă nu există evaluări
- Anexo-A NUESTRO PADRE CREADOR TÚPAC AMARUDocument3 paginiAnexo-A NUESTRO PADRE CREADOR TÚPAC AMARUabrahanmauriciosahuaÎncă nu există evaluări
- BORRADOR PRACTICA 11 y 14Document8 paginiBORRADOR PRACTICA 11 y 14vanesaÎncă nu există evaluări
- School Brass 2Document13 paginiSchool Brass 2my domp100% (1)
- Modelo Kaizen para La Solución de Problemas - V5Document86 paginiModelo Kaizen para La Solución de Problemas - V5Edwin DZÎncă nu există evaluări
- Unidad 1 El Sentido de Aprender Sobre EticaDocument41 paginiUnidad 1 El Sentido de Aprender Sobre EticaAlejandro Lascurais67% (3)
- GLOBALIZACIÓNDocument2 paginiGLOBALIZACIÓNCRISTIAN ALBERTO PINEDA ROMEROÎncă nu există evaluări
- Tu CalificaciónDocument3 paginiTu CalificaciónJose Vasquez0% (2)
- Tarea 2 Ingenieria EconomicaDocument7 paginiTarea 2 Ingenieria EconomicaAlexander Merejildo Tomalá76% (17)
- Matematica Financiera Act 5 Y6Document14 paginiMatematica Financiera Act 5 Y6Viedma JennyÎncă nu există evaluări
- Sesión #03: Hábitos de Estudio Reto AcadémicoDocument9 paginiSesión #03: Hábitos de Estudio Reto AcadémicoPilar Carbajal AzaldeguiÎncă nu există evaluări
- SESIÓN 01 Bienvenida Año 2021Document7 paginiSESIÓN 01 Bienvenida Año 2021Maricita An CuBeÎncă nu există evaluări
- GuíaDocument38 paginiGuíaBrayan LedezmaÎncă nu există evaluări
- Manual AlumnoDocument18 paginiManual AlumnoAlejandro HerrerosÎncă nu există evaluări
- Cultura XincaDocument16 paginiCultura XincaMelissa RamirezÎncă nu există evaluări
- Res RTDocument38 paginiRes RTSusy AlvisÎncă nu există evaluări
- 38 TCP Procesos y Productos de Textil, Confección y Piel PDFDocument14 pagini38 TCP Procesos y Productos de Textil, Confección y Piel PDFIsabel Moran Areces100% (3)
- Contrato de EstudioDocument5 paginiContrato de EstudioRosher Heli75% (4)
- Ensayo Geologia de ColombiaDocument5 paginiEnsayo Geologia de ColombiaDiana QuintanaÎncă nu există evaluări