S-ar putea să vă placă și
- Eye Movements Clinical CorrelationDocument8 paginiEye Movements Clinical CorrelationAsena TuiketeiÎncă nu există evaluări
- Elimination Disorder: Presented by Saba AhmedDocument49 paginiElimination Disorder: Presented by Saba AhmedRana Ahmad Gulraiz100% (1)
- Brain Edema & It's ManagementDocument26 paginiBrain Edema & It's ManagementMohammed FareedÎncă nu există evaluări
- ChiroandosteoDocument6 paginiChiroandosteoapi-3701230Încă nu există evaluări
- Cluster Headache - MedlinePlus Medical EncyclopediaDocument3 paginiCluster Headache - MedlinePlus Medical EncyclopediaUtpal KumarÎncă nu există evaluări
- Epigenetics of EndometriosisDocument21 paginiEpigenetics of EndometriosisirwinaÎncă nu există evaluări
- Hip Fracture of Proximal Femur SOAP COMPLETEDocument10 paginiHip Fracture of Proximal Femur SOAP COMPLETEEj TuliaoÎncă nu există evaluări
- Axillary ArteryDocument16 paginiAxillary ArteryHarsha MudigoudaÎncă nu există evaluări
- Toxins and Parasites: Rose Jean S. MontibanoDocument35 paginiToxins and Parasites: Rose Jean S. Montibanorose singuayÎncă nu există evaluări
- ClaustrophobiaDocument6 paginiClaustrophobiaZulhaime AyobÎncă nu există evaluări
- Cerebrospinal Liquor and Its CirculationDocument13 paginiCerebrospinal Liquor and Its CirculationusergrÎncă nu există evaluări
- Nuclear Physics A Level RevisionDocument32 paginiNuclear Physics A Level Revisionsupniggas80% (5)
- Eating DisordersDocument9 paginiEating DisordersFatin Nurina Mohd NazemiÎncă nu există evaluări
- Health, Stress, and CopingDocument43 paginiHealth, Stress, and CopingKar Gayee100% (1)
- Efferent Visual System (Ocular Motor Pathways)Document38 paginiEfferent Visual System (Ocular Motor Pathways)riskhapangestika100% (1)
- Dysphagia SurgicalDocument75 paginiDysphagia Surgicalian ismail100% (1)
- Chiropractic: An Introduction: Key PointsDocument6 paginiChiropractic: An Introduction: Key Pointsblackhole93Încă nu există evaluări
- Abnormal Midwifery: by Gladys M. BSN, KRCHNDocument352 paginiAbnormal Midwifery: by Gladys M. BSN, KRCHNMercy KeruboÎncă nu există evaluări
- Mock Board Exam in Microbiology/ Virology/Mycology/ParasitologyDocument12 paginiMock Board Exam in Microbiology/ Virology/Mycology/ParasitologyShera Heart Go100% (1)
- Central Nervous SystemDocument52 paginiCentral Nervous SystemRizki PerdanaÎncă nu există evaluări
- Chiropractics: General InformationDocument17 paginiChiropractics: General Informationapi-365631457Încă nu există evaluări
- Presented by DR Mohammed AtifDocument42 paginiPresented by DR Mohammed AtifUmer HussainÎncă nu există evaluări
- Cystic Fibrosis 17Document40 paginiCystic Fibrosis 17kholoud220Încă nu există evaluări
- Relaxation TechniquesDocument2 paginiRelaxation TechniquesFernando Williams GuibertÎncă nu există evaluări
- Animals Without Backbones1 PDFDocument31 paginiAnimals Without Backbones1 PDFSamKris Guerrero MalasagaÎncă nu există evaluări
- GCSE Business NotesDocument43 paginiGCSE Business NotesGadgetshop Station33% (3)
- Lupus Nephritis: Dr. Anchal Sahoo PRECEPTOR-Asso. Prof Dr. S.C. SinghDocument53 paginiLupus Nephritis: Dr. Anchal Sahoo PRECEPTOR-Asso. Prof Dr. S.C. Singhanchal sahooÎncă nu există evaluări
- Telomerase and AgingDocument10 paginiTelomerase and AgingPete WhiteÎncă nu există evaluări
- Cataracts: What Are They?Document3 paginiCataracts: What Are They?RSarkawyÎncă nu există evaluări
- Sexual and Asexual Reproduction Detailed Lesson PlanDocument9 paginiSexual and Asexual Reproduction Detailed Lesson PlanChristianJamesCapiz96% (25)
- Alzheimers-Disease-Genetics-Fact-Sheet 0Document8 paginiAlzheimers-Disease-Genetics-Fact-Sheet 0api-285676076Încă nu există evaluări
- Head & Neck - Deep Cervical Fascia &cervical PlexusDocument9 paginiHead & Neck - Deep Cervical Fascia &cervical PlexusKomang Tri Adi SuparwatiÎncă nu există evaluări
- Physics Mechanics Help BookletDocument88 paginiPhysics Mechanics Help Bookletdj7597100% (1)
- Biofloc TechnologyDocument53 paginiBiofloc Technologyjact_1100% (8)
- Grade 10 Daily Lesson PlanDocument3 paginiGrade 10 Daily Lesson PlanRod ReyesÎncă nu există evaluări
- Head Trauma: Adhitya Rahadi, M.DDocument50 paginiHead Trauma: Adhitya Rahadi, M.DNatasya Saraswati100% (1)
- 1-Basic Concepts and Theories of AgingDocument17 pagini1-Basic Concepts and Theories of AgingAaron KC SylvestreÎncă nu există evaluări
- Canas, Patricia Gan, Razel Ruelo, Vanessa Aquino, Camile Guillano, Christabelle BSN IiiDocument73 paginiCanas, Patricia Gan, Razel Ruelo, Vanessa Aquino, Camile Guillano, Christabelle BSN IiiGan BangÎncă nu există evaluări
- Fungal Evolution: Diversity, Taxonomy and Phylogeny of The FungiDocument37 paginiFungal Evolution: Diversity, Taxonomy and Phylogeny of The FungiPaula GeorgianaÎncă nu există evaluări
- Photosynthesis and Cellular RespirationDocument18 paginiPhotosynthesis and Cellular Respirationmaria genio100% (2)
- Pituitary GlandDocument22 paginiPituitary GlandSara Musavi100% (2)
- Cytogenetics Transes Topic 16Document5 paginiCytogenetics Transes Topic 16Dianne NolascoÎncă nu există evaluări
- Administering Medication Via A Small-Volume NebulizerDocument2 paginiAdministering Medication Via A Small-Volume NebulizerJerilee SoCute WattsÎncă nu există evaluări
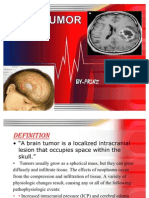
- PRADEEP'S - Brain TumorDocument32 paginiPRADEEP'S - Brain TumorPRADEEPÎncă nu există evaluări
- Water and Electrolyte ImbalanceDocument17 paginiWater and Electrolyte Imbalancemhairat6043100% (1)
- RicketsDocument53 paginiRicketsajitar20030% (1)
- Werner SyndromeDocument12 paginiWerner SyndromeCharles IppolitoÎncă nu există evaluări
- Personalized Cell Therapy: Tackling Some of The Greatest Unmet Needs in HealthcareDocument10 paginiPersonalized Cell Therapy: Tackling Some of The Greatest Unmet Needs in HealthcareKevin ParkerÎncă nu există evaluări
- ChiropracticDocument4 paginiChiropracticYe Myint100% (1)
- AgeLOC Science Year in Review 2010Document16 paginiAgeLOC Science Year in Review 2010dmatei101Încă nu există evaluări
- Neurochemical Cascade of ConcussionDocument16 paginiNeurochemical Cascade of ConcussionJoan Choi100% (1)
- Physiology of PainDocument35 paginiPhysiology of PainJauhar Firdaus50% (2)
- Immunity: Immunity Is The Balanced State ofDocument1 paginăImmunity: Immunity Is The Balanced State ofCarolina GasperÎncă nu există evaluări
- Introduction To Lymphoid Tissue & Immune System & Histology of Lymph Node & Thymus PDFDocument12 paginiIntroduction To Lymphoid Tissue & Immune System & Histology of Lymph Node & Thymus PDFhassam qaziÎncă nu există evaluări
- The Lymphatic System SummaryDocument3 paginiThe Lymphatic System SummaryVasile100% (1)
- Hip FractureDocument4 paginiHip Fracturemariafub100% (1)
- Types of CancerDocument7 paginiTypes of CancerRegina SABSÎncă nu există evaluări
- A StyeDocument12 paginiA StyeNaomi Cyden YapÎncă nu există evaluări
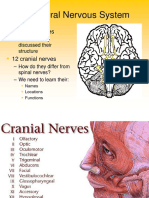
- Figure 1: Inferior View of The Brain and Brain Stem Showing Cranial NervesDocument40 paginiFigure 1: Inferior View of The Brain and Brain Stem Showing Cranial NervesVishnu Karunakaran0% (1)
- Soil Transmitted Helminth Infections: The Nature, Causes and Burden of The ConditionDocument79 paginiSoil Transmitted Helminth Infections: The Nature, Causes and Burden of The ConditionYokaiCO100% (1)
- Constipation: DefinitionDocument5 paginiConstipation: DefinitionSanthosh KumarÎncă nu există evaluări
- Gastritis: Department of Gastroenterology General Hospital of Ningxia Medical University Si Cen MDDocument82 paginiGastritis: Department of Gastroenterology General Hospital of Ningxia Medical University Si Cen MDAvi Themessy100% (1)
- Histo - Pituitary GlandDocument22 paginiHisto - Pituitary GlandRessam NazirÎncă nu există evaluări
- Drug Study: Submitted To: Mr. Blas Blasil A. PurgananDocument7 paginiDrug Study: Submitted To: Mr. Blas Blasil A. Purganancharanjit kaurÎncă nu există evaluări
- NCP 2 Impaired Skin Integrity EDITEDDocument2 paginiNCP 2 Impaired Skin Integrity EDITEDVincent HermanoÎncă nu există evaluări
- PHG 413 - Vitamin C b1 b2 b3Document56 paginiPHG 413 - Vitamin C b1 b2 b3Avicena RasyidÎncă nu există evaluări
- Fluorouracil PDFDocument8 paginiFluorouracil PDFKristine Dwi PuspitasariÎncă nu există evaluări
- 4 - CysticercosisDocument18 pagini4 - CysticercosisPhan Cảnh TrìnhÎncă nu există evaluări
- Fluids & Electrolytes Disorders: Ariano Marcos State UniversityDocument13 paginiFluids & Electrolytes Disorders: Ariano Marcos State UniversityClements galiza100% (1)
- Myasthenia Gravis: An Autoimmune Neurologic DisorderDocument16 paginiMyasthenia Gravis: An Autoimmune Neurologic DisorderHibba NasserÎncă nu există evaluări
- Psychiatric Effects of ThyroidDocument9 paginiPsychiatric Effects of ThyroidJosetta WhitneyÎncă nu există evaluări
- By: Noor Majeed RehaniDocument23 paginiBy: Noor Majeed RehaniMihaela Toma0% (1)
- The Tenses in English GrammarDocument6 paginiThe Tenses in English GrammarRSarkawyÎncă nu există evaluări
- 7096 IAL June 2014 Timetable - WEB v2Document11 pagini7096 IAL June 2014 Timetable - WEB v2Nithoo NishÎncă nu există evaluări
- TelomereDocument38 paginiTelomereRSarkawyÎncă nu există evaluări
- DiseasesDocument21 paginiDiseasesRSarkawy100% (1)
- 6PH01 01 Pef 20130815Document66 pagini6PH01 01 Pef 20130815RSarkawyÎncă nu există evaluări
- Physics Matter Help BookletDocument18 paginiPhysics Matter Help Bookletdj7597Încă nu există evaluări
- Society and Sex RolesDocument5 paginiSociety and Sex RolesRSarkawyÎncă nu există evaluări
- Advice Letters PDFDocument5 paginiAdvice Letters PDFRSarkawyÎncă nu există evaluări
- Edexcel As BiologyDocument1 paginăEdexcel As BiologyRSarkawy100% (1)
- Complete Advanced Level Mathematics - Pure MathematicsDocument512 paginiComplete Advanced Level Mathematics - Pure MathematicsRSarkawy86% (14)
- Unit 2Document14 paginiUnit 2RSarkawyÎncă nu există evaluări
- 8BI01 GCE08 Biology Rep 20090813 US021143 Unit 3Document12 pagini8BI01 GCE08 Biology Rep 20090813 US021143 Unit 3nourhanjÎncă nu există evaluări
- Freindly Letters - Descriptive PDFDocument11 paginiFreindly Letters - Descriptive PDFRSarkawyÎncă nu există evaluări
- Form Design 1 & 2Document1 paginăForm Design 1 & 2RSarkawyÎncă nu există evaluări
- Biology RevisionDocument3 paginiBiology RevisionMiguel HoffmannÎncă nu există evaluări
- Edexcel GCE Biology - AS Unit 3: Heart Rate / BPMDocument1 paginăEdexcel GCE Biology - AS Unit 3: Heart Rate / BPMRSarkawyÎncă nu există evaluări
- June 2012 OLs Grade ThresholdsDocument30 paginiJune 2012 OLs Grade ThresholdsRSarkawyÎncă nu există evaluări
- Behaviour in AnimalsDocument14 paginiBehaviour in AnimalsRSarkawyÎncă nu există evaluări
- 4BI0 1B Que 20130109Document28 pagini4BI0 1B Que 20130109Sharif MahmudÎncă nu există evaluări
- Edexcel Topic 2 Energy ChangesDocument4 paginiEdexcel Topic 2 Energy ChangesSan SiddzÎncă nu există evaluări
- CH 5 Test Yourself Q & ADocument1 paginăCH 5 Test Yourself Q & ARSarkawyÎncă nu există evaluări
- CH 23 Notes P2Document1 paginăCH 23 Notes P2RSarkawyÎncă nu există evaluări
- Answers On Next PageDocument5 paginiAnswers On Next PageRSarkawyÎncă nu există evaluări
- IGCSE Business Studies Revision GuideDocument12 paginiIGCSE Business Studies Revision GuideRSarkawy50% (4)
- What Causes Cystic Fibrosis1Document8 paginiWhat Causes Cystic Fibrosis1RSarkawyÎncă nu există evaluări
- Animal Transport SystemDocument24 paginiAnimal Transport SystemCamille TolentinoÎncă nu există evaluări
- Bla Bla Bla New 2014Document11 paginiBla Bla Bla New 2014Georgiana GeoÎncă nu există evaluări
- Bio1227 Microbiology and Parasitology Department of Biology Institute of Arts and Sciences Far Eastern UniversityDocument16 paginiBio1227 Microbiology and Parasitology Department of Biology Institute of Arts and Sciences Far Eastern UniversityDannielle EvangelistaÎncă nu există evaluări
- Daftar Pustaka PembahasanDocument2 paginiDaftar Pustaka PembahasanMaryam Fathiya RinthaniÎncă nu există evaluări
- Breast Cancer in The Views of Foods, Herbs & VitaminsDocument66 paginiBreast Cancer in The Views of Foods, Herbs & VitaminsKyle J. NortonÎncă nu există evaluări
- Discovering PhotosynthesisDocument3 paginiDiscovering PhotosynthesisEarl CaridadÎncă nu există evaluări
- Company. PP 135-152.: ReferencesDocument10 paginiCompany. PP 135-152.: ReferencesGlascow universityÎncă nu există evaluări
- As91603 Plant and Animal ResponsesDocument2 paginiAs91603 Plant and Animal Responsesapi-323107386Încă nu există evaluări
- Virus Introduction of Virus: Martinus Beijerinck Discovered The Tobacco Mosaic Virus (TMV) From TheDocument5 paginiVirus Introduction of Virus: Martinus Beijerinck Discovered The Tobacco Mosaic Virus (TMV) From Therafiul muttakimÎncă nu există evaluări
- NOTES - Module 2. Developmental and Reproductive BiologyDocument3 paginiNOTES - Module 2. Developmental and Reproductive BiologyTrxsee RoxasÎncă nu există evaluări
- Thesis PCRDocument8 paginiThesis PCRgjfs5mtv100% (2)
- Yr 8 WK 2 Bio NoteDocument3 paginiYr 8 WK 2 Bio Notesedrick ocheÎncă nu există evaluări
- SerotoninaDocument12 paginiSerotoninajodieÎncă nu există evaluări
- Blood Type Genetics ProtocolDocument5 paginiBlood Type Genetics ProtocolmakisgeraÎncă nu există evaluări
- Worksheet 9.1: Chapter 9: Plant Science - Fifteen Summary FactsDocument2 paginiWorksheet 9.1: Chapter 9: Plant Science - Fifteen Summary FactsRabia RafiqueÎncă nu există evaluări
- General Biology: Osmosis LabDocument9 paginiGeneral Biology: Osmosis Labapi-334420312Încă nu există evaluări
- Staats 2005Document14 paginiStaats 2005lacisagÎncă nu există evaluări
- The Evolution of Bizarre Structures' in Dinosaurs: Biomechanics, Sexual Selection, Social Selection or Species Recognition?Document15 paginiThe Evolution of Bizarre Structures' in Dinosaurs: Biomechanics, Sexual Selection, Social Selection or Species Recognition?Manuel PérezÎncă nu există evaluări
- Module 6 Science 10Document20 paginiModule 6 Science 10bhrayancacheroÎncă nu există evaluări
- Anatomy Tubuh ManusiaDocument12 paginiAnatomy Tubuh ManusiaMuhammad YusufÎncă nu există evaluări
- Sagala (2011)Document8 paginiSagala (2011)Ahmad ZulfikarÎncă nu există evaluări