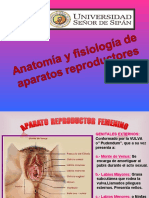
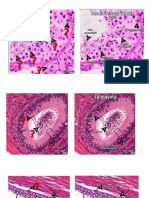
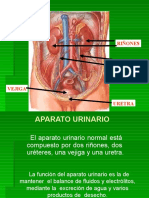
S-ar putea să vă placă și
- Mecanismos de La ApoptosisDocument6 paginiMecanismos de La ApoptosisMariela Alejandra Pinillos CuyunÎncă nu există evaluări
- Mecanismo de Patogenicidad BacterianaDocument3 paginiMecanismo de Patogenicidad BacterianaKeyla Vente VelascoÎncă nu există evaluări
- Sistema inmunitario: definición, componentes y funcionesDocument17 paginiSistema inmunitario: definición, componentes y funcionesCarlos Zanns CarpioÎncă nu există evaluări
- Células SanguíneasDocument8 paginiCélulas SanguíneasNadia ReyesÎncă nu există evaluări
- Complejo de Mayor HistocompatibilidadDocument7 paginiComplejo de Mayor HistocompatibilidadJonatán Eliseo Morales JeronimoÎncă nu există evaluări
- Tema 9 MicobacteriasDocument72 paginiTema 9 MicobacteriasJesus VilchezÎncă nu există evaluări
- Inmunidad Frente A Los TumoresDocument4 paginiInmunidad Frente A Los TumoresJacs SaulÎncă nu există evaluări
- Inmunidad frente a bacterias intracelulares y extracelularesDocument53 paginiInmunidad frente a bacterias intracelulares y extracelularesAndrea De los Reyes100% (1)
- ART - Revision1 Chancroide-2014 PDFDocument7 paginiART - Revision1 Chancroide-2014 PDFGAuReyesÎncă nu există evaluări
- Apuntes de Propedéutica de La Clínica 2024 PDFDocument113 paginiApuntes de Propedéutica de La Clínica 2024 PDFVanessa RamirezÎncă nu există evaluări
- Glosario de BioquimicaDocument8 paginiGlosario de Bioquimicaapi-259248355Încă nu există evaluări
- Complejo Mayor de HistocompatibilidadDocument23 paginiComplejo Mayor de HistocompatibilidadSandy Montoya100% (1)
- HEMATOPOYESISDocument6 paginiHEMATOPOYESISLaura Fernanda Díaz100% (1)
- Memoria InmunológicaDocument21 paginiMemoria InmunológicaGYadiraMiramontesHdz50% (2)
- Fisiología femenina: ciclo ovárico, hormonas y efectosDocument3 paginiFisiología femenina: ciclo ovárico, hormonas y efectosastrid viverosÎncă nu există evaluări
- Revisión Bibliográfica de Complejo Mayor de HistocompatibilidadDocument5 paginiRevisión Bibliográfica de Complejo Mayor de HistocompatibilidadOtoney Intriago100% (2)
- Sistema Inmune 2018 Tipeo 2Document34 paginiSistema Inmune 2018 Tipeo 2Belen Hernandez AguilarÎncă nu există evaluări
- Casos Clínicos Cocos Gram Positivos PDFDocument4 paginiCasos Clínicos Cocos Gram Positivos PDFAlisson CadenaÎncă nu există evaluări
- ELECTROFISIOLOGIADocument91 paginiELECTROFISIOLOGIAHeber ReyesÎncă nu există evaluări
- DiencefaloDocument33 paginiDiencefaloYvette Cabrera RaaÎncă nu există evaluări
- Membrana CelularDocument22 paginiMembrana CelularManuel Cespedes100% (1)
- Resistencia Del Organismo A La InfecciónDocument4 paginiResistencia Del Organismo A La InfecciónMayki QuintanaÎncă nu există evaluări
- Fagocitosis e InflamaciónDocument8 paginiFagocitosis e InflamaciónDouglasUrquillaÎncă nu există evaluări
- Inmuno TestDocument31 paginiInmuno TestSebas ArcosÎncă nu există evaluări
- Procesamiento Presentacion Antigenica PDFDocument28 paginiProcesamiento Presentacion Antigenica PDFRichard Bravo CéspedesÎncă nu există evaluări
- Moleculas de AdhesionDocument10 paginiMoleculas de AdhesionTefita XivitaÎncă nu există evaluări
- LPS y Receptores Toll (MAOG)Document13 paginiLPS y Receptores Toll (MAOG)miguel angel100% (3)
- ClostridiumDocument42 paginiClostridiumAaron Villarreal PereyraÎncă nu există evaluări
- Seminario de Celulas DendriticasDocument24 paginiSeminario de Celulas DendriticasAnonymous IuimQGqnmÎncă nu există evaluări
- Organos y Tejidos InmuneDocument31 paginiOrganos y Tejidos InmuneAlberto Guillermo Basurto MobarakÎncă nu există evaluări
- Resumen Inmunologia AcoDocument135 paginiResumen Inmunologia AcoManuel González AlayesÎncă nu există evaluări
- Replicacion de RubeolaDocument2 paginiReplicacion de RubeolaDaniela GomezÎncă nu există evaluări
- Inflamación y Reparación TisularDocument11 paginiInflamación y Reparación TisularBetty CarreñoÎncă nu există evaluări
- Inmunidad-Innata y AdquiridaDocument63 paginiInmunidad-Innata y Adquiridaliliana loyolaÎncă nu există evaluări
- Corynebacterium DiphtheriaeDocument11 paginiCorynebacterium DiphtheriaePaco HernandezÎncă nu există evaluări
- ParamyxoviridaeDocument4 paginiParamyxoviridaealexandra paez vargas - -Încă nu există evaluări
- Fisiología de Los Granulocitos.Document20 paginiFisiología de Los Granulocitos.medicinaens100% (3)
- Tercer Parcial PATOLOGÍADocument37 paginiTercer Parcial PATOLOGÍAVale Echavarri100% (1)
- Resumen BacteriologiaDocument15 paginiResumen BacteriologiajavierÎncă nu există evaluări
- Regulacion MolecularDocument30 paginiRegulacion MolecularAlexandra Aguilar0% (1)
- INFECCIONES ESTAFILOCÓCICAS: S. AUREUS Y COAGULASANEGATIVOSDocument17 paginiINFECCIONES ESTAFILOCÓCICAS: S. AUREUS Y COAGULASANEGATIVOSmichiiiÎncă nu există evaluări
- Uniones celulares: comunicantes, adherentes y oclusivasDocument3 paginiUniones celulares: comunicantes, adherentes y oclusivasAndreaÎncă nu există evaluări
- Barreras AnatómicasDocument7 paginiBarreras AnatómicasFay CerónÎncă nu există evaluări
- Uso y Abuso de Los AntibióticosDocument4 paginiUso y Abuso de Los AntibióticosElena ZarachoÎncă nu există evaluări
- Ciclo Reproductivo de Los VirusDocument7 paginiCiclo Reproductivo de Los VirusAndRee JaramIlloÎncă nu există evaluări
- 7.reparacion CelularDocument3 pagini7.reparacion CelularDaniel Gomez Torres100% (1)
- Trastornos Del Metabolismo Del Agua y Electrolitos Dr. Fredy Sandi LDocument10 paginiTrastornos Del Metabolismo Del Agua y Electrolitos Dr. Fredy Sandi LMachaca RonaldÎncă nu există evaluări
- Anatomía e Histología Del Sistema RenalDocument8 paginiAnatomía e Histología Del Sistema RenalDessiree PinkcessÎncă nu există evaluări
- Histo Sem 01 (Técnica Histológica)Document124 paginiHisto Sem 01 (Técnica Histológica)Celso NemerÎncă nu există evaluări
- Resumen Capitulo 12 Libro Genética en Medicina Thompson & ThompsonDocument8 paginiResumen Capitulo 12 Libro Genética en Medicina Thompson & ThompsonSamuelQuinde0% (1)
- Diapositivas de EstafilococosDocument33 paginiDiapositivas de EstafilococosMilagrOs Benites Reyes100% (1)
- Vacunas Bacterianas IDocument20 paginiVacunas Bacterianas IRUBEN JUNIOR LOPEZ AREVALOÎncă nu există evaluări
- Chlamydia TrachomatisDocument8 paginiChlamydia TrachomatisJUAN MANUEL CERON ALVARADOÎncă nu există evaluări
- Micosis Endemicas (Profundas o Sistemicas) - Cata (Autoguardado)Document9 paginiMicosis Endemicas (Profundas o Sistemicas) - Cata (Autoguardado)Isabella Lozano SantaÎncă nu există evaluări
- Parásitos Hemáticos y TisularesDocument5 paginiParásitos Hemáticos y TisularesHector Alejandro Salas GuzmanÎncă nu există evaluări
- Conceptos Básicos de La Respuesta Inmunitaria PDFDocument67 paginiConceptos Básicos de La Respuesta Inmunitaria PDFLuis Fernando Velasquez SantamariaÎncă nu există evaluări
- GneroneisseriaDocument36 paginiGneroneisseriamiriam ormeoÎncă nu există evaluări
- Casos Clínicos VirologíaDocument8 paginiCasos Clínicos VirologíaARELY MURIEL MORENOÎncă nu există evaluări
- Respuesta Inmune ViralDocument24 paginiRespuesta Inmune ViralJesus Andres MonroyÎncă nu există evaluări
- Receptores de las células T: estructura y función del TCRDocument19 paginiReceptores de las células T: estructura y función del TCRBrenda GonzalesÎncă nu există evaluări
- Cirugia General 3Document15 paginiCirugia General 3LeninReyesÎncă nu există evaluări
- Modelo de Fua - 010Document2 paginiModelo de Fua - 010LeninReyes0% (1)
- Cronograma en Vivo RM 2022Document1 paginăCronograma en Vivo RM 2022LeninReyesÎncă nu există evaluări
- MODELO DE FUA - 015 UnicoDocument2 paginiMODELO DE FUA - 015 UnicoLeninReyesÎncă nu există evaluări
- Modelo de Fua - 011Document2 paginiModelo de Fua - 011LeninReyesÎncă nu există evaluări
- Cartilla Promsa-Mcr ChorosDocument17 paginiCartilla Promsa-Mcr ChorosLeninReyesÎncă nu există evaluări
- Modelo de Fua - 075 UnicaDocument2 paginiModelo de Fua - 075 UnicaLeninReyes0% (1)
- REVISIÓN GINECOLÓGICA ANUAL: TOPE 1 AL AÑODocument2 paginiREVISIÓN GINECOLÓGICA ANUAL: TOPE 1 AL AÑOLeninReyes0% (1)
- El ConocimientoDocument20 paginiEl ConocimientoLeninReyesÎncă nu există evaluări
- MODELO DE FUA - 056 UnicoDocument2 paginiMODELO DE FUA - 056 UnicoLeninReyes100% (1)
- Modelo de Fua - 054 de I-1 Al 1-3Document2 paginiModelo de Fua - 054 de I-1 Al 1-3LeninReyesÎncă nu există evaluări
- Desarrollo Metodologico y Comprobacion de Hipotesis - Metodologia 6Document35 paginiDesarrollo Metodologico y Comprobacion de Hipotesis - Metodologia 6LeninReyesÎncă nu există evaluări
- Modelo de Fua - 022 PDFDocument2 paginiModelo de Fua - 022 PDFLeninReyesÎncă nu există evaluări
- Modelo de Fua - 018Document2 paginiModelo de Fua - 018LeninReyesÎncă nu există evaluări
- MODELO DE FUA - 071 UnicaDocument2 paginiMODELO DE FUA - 071 UnicaLeninReyes0% (1)
- Exposicion 26-50Document64 paginiExposicion 26-50LeninReyesÎncă nu există evaluări
- Vademecum FarmacologicoDocument459 paginiVademecum FarmacologicoJesus Alberto Rojas Castro100% (1)
- Cirugia General 3Document56 paginiCirugia General 3ANtonyMV0% (1)
- Presentacion Pediatria I - Enam 2014Document288 paginiPresentacion Pediatria I - Enam 2014LeninReyesÎncă nu există evaluări
- Anatomía 11ed-2019 PDFDocument30 paginiAnatomía 11ed-2019 PDFdanitza pilco100% (1)
- Lesión hipodensa frontoparietal derecha por isquemia cerebralDocument7 paginiLesión hipodensa frontoparietal derecha por isquemia cerebralLeninReyesÎncă nu există evaluări
- Examen de Pediatría ENAM 2014 Cap4Document301 paginiExamen de Pediatría ENAM 2014 Cap4LeninReyesÎncă nu există evaluări
- Anatomía IDocument49 paginiAnatomía ILeninReyesÎncă nu există evaluări
- NEUROLOGÍADocument7 paginiNEUROLOGÍALeninReyesÎncă nu există evaluări
- Cadena Epidemiológica Del DengueDocument2 paginiCadena Epidemiológica Del DengueLeninReyesÎncă nu există evaluări
- Consenso EPI 2017Document13 paginiConsenso EPI 2017JUAN FRANCISCOÎncă nu există evaluări
- Test Pediatria 01 04 Dr. GarciaDocument330 paginiTest Pediatria 01 04 Dr. GarciaLeninReyesÎncă nu există evaluări
- Examen de Pediatría ENAM 2014 Cap 5Document476 paginiExamen de Pediatría ENAM 2014 Cap 5LeninReyesÎncă nu există evaluări
- Examen de Pediatría ENAM Cap 6 11 08Document460 paginiExamen de Pediatría ENAM Cap 6 11 08LeninReyesÎncă nu există evaluări
- Curso de Preparación ENAM 2014 Cap3 Pediatria 13 05Document335 paginiCurso de Preparación ENAM 2014 Cap3 Pediatria 13 05LeninReyesÎncă nu există evaluări
- Galeria 2 de La 8 Semana Al NacimientoDocument15 paginiGaleria 2 de La 8 Semana Al NacimientoJenniffer RamirezÎncă nu există evaluări
- Aparato Reproductor GreyDocument26 paginiAparato Reproductor GreyAndres HerediaÎncă nu există evaluări
- Sistema somatosensorial: receptores sensorialesDocument42 paginiSistema somatosensorial: receptores sensorialesDrakeantoniÎncă nu există evaluări
- Vasos Extraordinarios y Sus Areas de InfluenciaDocument1 paginăVasos Extraordinarios y Sus Areas de InfluenciaArelis del Rio100% (1)
- Arterias y Venas de Miembros InferioresDocument48 paginiArterias y Venas de Miembros InferioresADVA VIAZOÎncă nu există evaluări
- Syllabus Anatomia IiDocument24 paginiSyllabus Anatomia IiAlhelyÎncă nu există evaluări
- 03 - Alteraciones de La ConcienciaDocument34 pagini03 - Alteraciones de La ConcienciaAle ZambranoÎncă nu există evaluări
- Resolución Ejercicios PresencialesDocument21 paginiResolución Ejercicios PresencialesDAINNEÎncă nu există evaluări
- Embrio 2Document10 paginiEmbrio 2Anette Michelle Romo RenteríaÎncă nu există evaluări
- Cavidades Corporales Mesenterios y DiafragmaDocument51 paginiCavidades Corporales Mesenterios y Diafragmacaleb apaza aruquipa100% (1)
- AP. MasculinoDocument26 paginiAP. MasculinoAaron MendozaÎncă nu există evaluări
- 600658Document16 pagini600658Daira VázquezÎncă nu există evaluări
- 9.-Patologia SalivalDocument39 pagini9.-Patologia SalivalSilverTelloFernadezÎncă nu există evaluări
- Examen IBM 2015Document17 paginiExamen IBM 2015Pedro Farro AbadÎncă nu există evaluări
- Aparato RespiratorioDocument9 paginiAparato RespiratorioNicolas MarunÎncă nu există evaluări
- Cuestionario Fisio - Gastro - UsmpDocument2 paginiCuestionario Fisio - Gastro - UsmpMaria Pia BeltranÎncă nu există evaluări
- Craneo y TroncoDocument9 paginiCraneo y TroncoChris NorÎncă nu există evaluări
- Clase EFMDocument1 paginăClase EFMFäbiänä Hernändez0% (1)
- Histología-Sistema ReproductorDocument21 paginiHistología-Sistema ReproductorDana Jurado MayorgaÎncă nu există evaluări
- TALLER Grado 8° D Reproduccion HUMANADocument3 paginiTALLER Grado 8° D Reproduccion HUMANASANTIAGO CARDONA100% (2)
- Anatomia y Fisiologia de Los RumiantesDocument37 paginiAnatomia y Fisiologia de Los Rumiantesdaddy rey0% (1)
- EmbrioDocument44 paginiEmbrioVladimir Vences GamaÎncă nu există evaluări
- Biomecanica-fisiologia-Anatomia Columna Lumbar PelvisDocument43 paginiBiomecanica-fisiologia-Anatomia Columna Lumbar PelvisCarlos Alberto Alvarez CortesÎncă nu există evaluări
- Reporte 2 AnatomiaDocument3 paginiReporte 2 AnatomiaVanezZiitaTramce100% (1)
- Medicina - Fisiologia. Líquidos Corporales 2007Document59 paginiMedicina - Fisiologia. Líquidos Corporales 2007DANIEL ALEJANDROÎncă nu există evaluări
- Tema 3.6. INFLAMACIÓN (Autoguardado)Document82 paginiTema 3.6. INFLAMACIÓN (Autoguardado)Eduardo Teran MontalvoÎncă nu există evaluări
- Anatomia Del Riñon y Vias UrinariasDocument95 paginiAnatomia Del Riñon y Vias UrinariasBeatriz Milagros PeveÎncă nu există evaluări
- Aparato Digestivo TextoDocument5 paginiAparato Digestivo TextoLucy Encalada NeiraÎncă nu există evaluări
- Fundamentos Biológicos Del Comportamiento HumanoDocument14 paginiFundamentos Biológicos Del Comportamiento HumanoSonia L. ValleÎncă nu există evaluări
- Afasia de Conduccion (Recovered 1)Document13 paginiAfasia de Conduccion (Recovered 1)Luis PadillaÎncă nu există evaluări