S-ar putea să vă placă și
- NIF A-5 Normas Del ActivoDocument1 paginăNIF A-5 Normas Del ActivoHugo RoqueÎncă nu există evaluări
- Boletin 0116Document13 paginiBoletin 0116Lucas FernandezÎncă nu există evaluări
- Paper DockDocument71 paginiPaper DockHugo RoqueÎncă nu există evaluări
- 518 2013 10 28 Ec - Cerradas PDFDocument25 pagini518 2013 10 28 Ec - Cerradas PDFAlfred S-aÎncă nu există evaluări
- DDD293 PDFDocument127 paginiDDD293 PDFKety Olortegui MelgarejoÎncă nu există evaluări
- Ev Sumativa 5am1hpDocument1 paginăEv Sumativa 5am1hpHugo RoqueÎncă nu există evaluări
- Infografia 3 Codigo de EticaDocument1 paginăInfografia 3 Codigo de EticaMarycela Osorio100% (1)
- Teoria Macroeconomica I Problemas y Ejer PDFDocument79 paginiTeoria Macroeconomica I Problemas y Ejer PDFJose Humberto Meneses Gonzales40% (5)
- Ejercicios Tema 3 Modelo Is LDocument2 paginiEjercicios Tema 3 Modelo Is LBrayan Smith Masias Ojeda50% (2)
- Set de Ejercicios Curso Introd. MacroDocument24 paginiSet de Ejercicios Curso Introd. Macroglasys16100% (3)
- 518 2013 10 28 Ec - Cerradas PDFDocument25 pagini518 2013 10 28 Ec - Cerradas PDFAlfred S-aÎncă nu există evaluări
- Macroeconomía Modelo Is-LmDocument40 paginiMacroeconomía Modelo Is-LmJoselyn Sharon100% (1)
- 518 2013 10 28 Ec - Cerradas PDFDocument25 pagini518 2013 10 28 Ec - Cerradas PDFAlfred S-aÎncă nu există evaluări
- Mercado de TrabajoDocument10 paginiMercado de TrabajoHugo RoqueÎncă nu există evaluări
- T M F K M: Luis Orduna DíezDocument18 paginiT M F K M: Luis Orduna DíezHugo RoqueÎncă nu există evaluări
- Macroeconomía Modelo Is-LmDocument40 paginiMacroeconomía Modelo Is-LmJoselyn Sharon100% (1)
- Preferencia Por Liquidez Como Comportamiento Frente Al RiesgoDocument14 paginiPreferencia Por Liquidez Como Comportamiento Frente Al Riesgoriemma100% (14)
- Ecuacin Slutsky EjercicioDocument24 paginiEcuacin Slutsky EjercicioMonica Carolina Retana LazoÎncă nu există evaluări
- 1517 2850 1 SMDocument9 pagini1517 2850 1 SMHugo RoqueÎncă nu există evaluări
- Tema 1 PDFDocument56 paginiTema 1 PDFmariofrrÎncă nu există evaluări
- Dialnet MetodosMatematicosYComputacionalesEnMacroeconomia 6876056 PDFDocument6 paginiDialnet MetodosMatematicosYComputacionalesEnMacroeconomia 6876056 PDFHugo RoqueÎncă nu există evaluări
- Explicacion Del Consumo Segun SmithiesDocument23 paginiExplicacion Del Consumo Segun SmithiesHugo RoqueÎncă nu există evaluări
- MACROECONOMIA I - Resumen Dornbusch y FischerDocument121 paginiMACROECONOMIA I - Resumen Dornbusch y FischerDavid Alejandro Gutierrez Silva58% (19)
- Teoria General KeynesDocument35 paginiTeoria General KeynesAlisson EspitiaTÎncă nu există evaluări
- Sugerencias Técnicas para La Elección de BombaDocument1 paginăSugerencias Técnicas para La Elección de BombaHugo RoqueÎncă nu există evaluări
- Tecnicas Reforzamiento Estructuras Construidas Concreto Deficiencias EstructuralesDocument31 paginiTecnicas Reforzamiento Estructuras Construidas Concreto Deficiencias EstructuralesYonathan Yesghar Pacheco CervantesÎncă nu există evaluări
- Bases Del TorneoDocument1 paginăBases Del TorneoHugo RoqueÎncă nu există evaluări
- 7590.66.59.2.acometidas en Media TensionDocument12 pagini7590.66.59.2.acometidas en Media Tensionmarlon_agzÎncă nu există evaluări
- Tecnicas Reforzamiento Estructuras Construidas Concreto Deficiencias EstructuralesDocument31 paginiTecnicas Reforzamiento Estructuras Construidas Concreto Deficiencias EstructuralesYonathan Yesghar Pacheco CervantesÎncă nu există evaluări
- Humedad PDFDocument70 paginiHumedad PDFMILAGROS MLTÎncă nu există evaluări
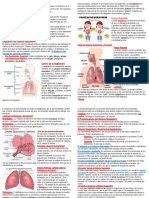
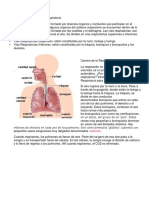
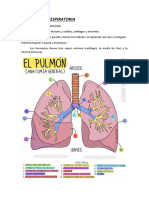
- Triptico Sistema Respiratorio 130130192924 Phpapp02Document2 paginiTriptico Sistema Respiratorio 130130192924 Phpapp02एलेक्स विगोÎncă nu există evaluări
- Examen 02 de Ciencia y Ambiente para Tercero de Primaria PDFDocument2 paginiExamen 02 de Ciencia y Ambiente para Tercero de Primaria PDFLinda Melendez0% (1)
- Cuadro de SistemasDocument2 paginiCuadro de SistemasLorena ArrietaÎncă nu există evaluări
- Sindrome de Distres Respiratorio Agudo Abordaje Basado en EvidenciaDocument16 paginiSindrome de Distres Respiratorio Agudo Abordaje Basado en Evidenciareinaldo valdes iznagaÎncă nu există evaluări
- Oxidaciones Biologicas MD RoxDocument49 paginiOxidaciones Biologicas MD RoxDAFNE ROSSY CABRERA DEZAÎncă nu există evaluări
- Manual Básico de VentilaciónDocument35 paginiManual Básico de Ventilaciónsiul100% (1)
- Fermentacion LacticaDocument5 paginiFermentacion LacticaNaturalness CosmeticsÎncă nu există evaluări
- HIPOXIADocument12 paginiHIPOXIAMARGIE RINCON TRIANAÎncă nu există evaluări
- Lab Espirometria IntegradoDocument9 paginiLab Espirometria IntegradoJUAN SEBASTIAN VARGAS SANCHEZÎncă nu există evaluări
- RespiratorioDocument17 paginiRespiratorioIvonne CMÎncă nu există evaluări
- Fisiología II RespiratorioDocument26 paginiFisiología II RespiratorioDaniel EspindolaÎncă nu există evaluări
- La Respiración de Los Seres VivosDocument3 paginiLa Respiración de Los Seres VivosAntonio TelloÎncă nu există evaluări
- Técnica de Respiración ProfundaDocument1 paginăTécnica de Respiración ProfundaMiguel AngelÎncă nu există evaluări
- Trabajo Practico Sistema RespiratorioDocument2 paginiTrabajo Practico Sistema RespiratorioVictoria DangeliÎncă nu există evaluări
- Clase 7. Insuficiencia Respiratoria AgudaDocument48 paginiClase 7. Insuficiencia Respiratoria AgudaYesiVZÎncă nu există evaluări
- BC 184 DDocument5 paginiBC 184 DDiego Enrique Venegas TipianÎncă nu există evaluări
- Cuidados de Enfermería en El Paciente Intubado AdultoDocument41 paginiCuidados de Enfermería en El Paciente Intubado AdultoDetallitos AnyÎncă nu există evaluări
- Clase 14 Principales Examenes Auxiliares Del Aparato Respiratorio en NeumologiaDocument9 paginiClase 14 Principales Examenes Auxiliares Del Aparato Respiratorio en NeumologiaMarc FloresmÎncă nu există evaluări
- Explicación Del Sistema RespiratorioDocument1 paginăExplicación Del Sistema Respiratoriooficial MisNotasBrevesÎncă nu există evaluări
- Manual Asistencia Ventilatoria PDFDocument67 paginiManual Asistencia Ventilatoria PDFAnonymous QC9PE2ZÎncă nu există evaluări
- OxigenoterapiaDocument5 paginiOxigenoterapiaMarco Flores MarceloÎncă nu există evaluări
- Sindromes Respiratorios 3Document16 paginiSindromes Respiratorios 3Leidy Yesenia Chura CallohuancaÎncă nu există evaluări
- Seminario 10 RespiratorioDocument9 paginiSeminario 10 RespiratorioPaulina FuentesÎncă nu există evaluări
- Fisiopatología Respiratoria Sem 04Document60 paginiFisiopatología Respiratoria Sem 04hemer hadyn calderon alvites0% (1)
- Free Aparato RespiratorioDocument10 paginiFree Aparato RespiratorioMarta Plata a veraÎncă nu există evaluări
- Transporte Electrónico y Fosforilación OxidativaDocument6 paginiTransporte Electrónico y Fosforilación OxidativaAbdiel SandrezÎncă nu există evaluări
- RESPIDocument8 paginiRESPIAmpa Ro DriguezÎncă nu există evaluări
- Crucigrama VeterianariaDocument2 paginiCrucigrama VeterianariaJenny HernandezÎncă nu există evaluări
- Plan de Cuidados - GRUPODocument7 paginiPlan de Cuidados - GRUPOAnonymous Bf1VqHPacÎncă nu există evaluări
- Modulo de Vantilacion Mecanica - CUIDADOS DE ENFERMERIA AL PACIENTE CON SOPORTE VENTILATORIO 2Document27 paginiModulo de Vantilacion Mecanica - CUIDADOS DE ENFERMERIA AL PACIENTE CON SOPORTE VENTILATORIO 2George Ruydiaz Ebratt100% (1)