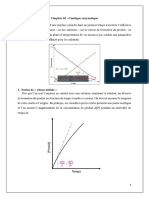
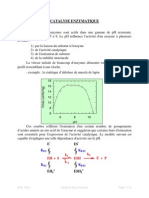
S-ar putea să vă placă și
- Mesure de Lactivité EnzymatiqueDocument23 paginiMesure de Lactivité EnzymatiqueIssam mairfatÎncă nu există evaluări
- Cinetique Enzymatique-1 PDFDocument15 paginiCinetique Enzymatique-1 PDFTCHEMMIK NazihaÎncă nu există evaluări
- 3a - Cinetique EnzymatiqueDocument8 pagini3a - Cinetique EnzymatiqueSameh Khemir100% (2)
- Biochimie1an-Cinetique Enzymatique2020feraga PDFDocument4 paginiBiochimie1an-Cinetique Enzymatique2020feraga PDFAsmaÎncă nu există evaluări
- Cinetique Enzyme 2 PDFDocument13 paginiCinetique Enzyme 2 PDFMer OuaneÎncă nu există evaluări
- Cours Enzymologie 2022Document86 paginiCours Enzymologie 2022Adja Aïta BaÎncă nu există evaluări
- TD BioreacteurDocument48 paginiTD Bioreacteurlola benr83% (6)
- Chapitre 4 Cinétique Et InhibitionDocument14 paginiChapitre 4 Cinétique Et Inhibitionkgu yryrtyÎncă nu există evaluări
- Cinétique EnzymatiqueDocument4 paginiCinétique EnzymatiqueAÎncă nu există evaluări
- Cours enz-PI-1Document61 paginiCours enz-PI-1antaafall464Încă nu există evaluări
- 10.03.2016 Les EnzymesDocument14 pagini10.03.2016 Les EnzymesEaÎncă nu există evaluări
- ENZYMOLOGIE II.2021pptDocument59 paginiENZYMOLOGIE II.2021pptKaren DominiqueÎncă nu există evaluări
- Biochimie1an-Cinetique Enzymatique2020feragaDocument27 paginiBiochimie1an-Cinetique Enzymatique2020feragaImene Aoun SeghirÎncă nu există evaluări
- Enzymologie TP Invertase PolycopieDocument14 paginiEnzymologie TP Invertase Polycopieiliass lahmassÎncă nu există evaluări
- Cours Cinétique Enzymatique. 21docxDocument7 paginiCours Cinétique Enzymatique. 21docxKEBIR ZAHRAÎncă nu există evaluări
- Kira DriveDocument7 paginiKira DriveManon righiniÎncă nu există evaluări
- Chapitre 02 Cinétique EnzymatiqueDocument5 paginiChapitre 02 Cinétique Enzymatiqueamina44Încă nu există evaluări
- La Cinétique EnzymatiqueDocument6 paginiLa Cinétique EnzymatiqueTCHEMMIK NazihaÎncă nu există evaluări
- TD1 CorrigeDocument12 paginiTD1 CorrigeLinda KoundziÎncă nu există evaluări
- Effecteur EnzymatiqueDocument6 paginiEffecteur EnzymatiqueAÎncă nu există evaluări
- 2 Enzym BasesDocument2 pagini2 Enzym BasesKhyarhoum Brahim100% (1)
- Cours equation henri michaelis menten briggs haldane constante catalytique vmax km lineweaver Burk Dixon Hanes Woolf Enzymologie Enseignement et recherche Biochimie Emmanuel Jaspard Universite Angers biochimej.pdfDocument13 paginiCours equation henri michaelis menten briggs haldane constante catalytique vmax km lineweaver Burk Dixon Hanes Woolf Enzymologie Enseignement et recherche Biochimie Emmanuel Jaspard Universite Angers biochimej.pdfTCHEMMIK NazihaÎncă nu există evaluări
- Voir Un Cours Sur Les Paramètres Cinétiques Des Réactions Ctalysées Par Les EnzymesDocument4 paginiVoir Un Cours Sur Les Paramètres Cinétiques Des Réactions Ctalysées Par Les EnzymesLAMINE DEGUISTARÎncă nu există evaluări
- Biochimie Approfondie TD 4Document4 paginiBiochimie Approfondie TD 4Alhassane BahÎncă nu există evaluări
- CH 8-EnzymesDocument44 paginiCH 8-Enzymesyaya camaraÎncă nu există evaluări
- Chap 11 CDocument4 paginiChap 11 CStachis Madiamba100% (1)
- AGB S2 M6.3 Enzymologie CRS BouslamtiDocument39 paginiAGB S2 M6.3 Enzymologie CRS BouslamtiLAMINE DEGUISTARÎncă nu există evaluări
- Cinetique Des Enzymes 2Document26 paginiCinetique Des Enzymes 2ZG NourelhoudaÎncă nu există evaluări
- 2 - Introduction A L - Enzymologie PDFDocument9 pagini2 - Introduction A L - Enzymologie PDFGabrielle NnomoÎncă nu există evaluări
- Génie Enzymologique - Asmaa BenjrinijaDocument19 paginiGénie Enzymologique - Asmaa BenjrinijaLEMRAHRI kaheliÎncă nu există evaluări
- Enzymologie TP Invertase PolycopieDocument16 paginiEnzymologie TP Invertase PolycopieEl Ibtihaal33% (3)
- 2 лекция ферментыDocument10 pagini2 лекция ферментыE BÎncă nu există evaluări
- Enzymologie Et BioénergétiqueDocument15 paginiEnzymologie Et BioénergétiqueZorba ZorbaÎncă nu există evaluări
- Exercices Diriges Enzymologie 2011 2012Document8 paginiExercices Diriges Enzymologie 2011 2012alphonse198867% (3)
- Cours de Cinétique ChimiqueDocument6 paginiCours de Cinétique Chimiqueilyes fellagÎncă nu există evaluări
- Toussaint Bertrand EnzyDocument85 paginiToussaint Bertrand EnzyZaki KhelifaÎncă nu există evaluări
- EnzymologieDocument12 paginiEnzymologieBasmã AlilecheÎncă nu există evaluări
- 4 - Catalyse Enzymatique PDFDocument14 pagini4 - Catalyse Enzymatique PDFPrince Abdoo100% (1)
- Corrigé TD MPC - GMSI-MRMI-EX 3-4-5-6-7-8Document11 paginiCorrigé TD MPC - GMSI-MRMI-EX 3-4-5-6-7-8Mohammed El azhariÎncă nu există evaluări
- Chapitre 5Document7 paginiChapitre 5maryam.dellalÎncă nu există evaluări
- CinetiqueDocument25 paginiCinetiqueOussama El BouadiÎncă nu există evaluări
- Vanhaverbeke Cecile p02 PDFDocument33 paginiVanhaverbeke Cecile p02 PDFMourad Makhlouf100% (1)
- Recueil D Exercices EnzymesDocument6 paginiRecueil D Exercices EnzymesStephanie TollenaereÎncă nu există evaluări
- Chapitre Vi Cinetique Des Reactions ComposeesDocument5 paginiChapitre Vi Cinetique Des Reactions ComposeesMidi SthÎncă nu există evaluări
- Cours M2 PINGPONGDocument8 paginiCours M2 PINGPONGAllah Beurem BonheurÎncă nu există evaluări
- 4e DS1 2010 2011Document4 pagini4e DS1 2010 2011fethi10% (1)
- Polycope TP Enzymologie - 2021Document13 paginiPolycope TP Enzymologie - 2021a.elabbassiÎncă nu există evaluări
- TD de Biochimie MetaboliqueDocument30 paginiTD de Biochimie MetaboliqueKnox ChaliÎncă nu există evaluări
- ΔG ' conditions standard 1 M: Voir les énoncés des exercicesDocument9 paginiΔG ' conditions standard 1 M: Voir les énoncés des exercicesLAMINE DEGUISTAR100% (7)
- Correction TD 2Document6 paginiCorrection TD 2Linda KoundziÎncă nu există evaluări
- EMD M1 Ailim 2019 Bioreacteur-Sol - TextmarkDocument3 paginiEMD M1 Ailim 2019 Bioreacteur-Sol - TextmarkOmairÎncă nu există evaluări
- TD Série 2 EnzymologieDocument3 paginiTD Série 2 EnzymologieAzzeddine KhartaÎncă nu există evaluări
- Partie 2 Enzymo FEV 12Document49 paginiPartie 2 Enzymo FEV 12Limoune AliÎncă nu există evaluări
- Introduction EnzymatologieDocument9 paginiIntroduction Enzymatologiefiras7Încă nu există evaluări
- 3 Chapitre II EnzymesDocument62 pagini3 Chapitre II EnzymesNour El HoudaÎncă nu există evaluări
- Cine 4Document4 paginiCine 4AliAliÎncă nu există evaluări
- Cinétique Enzymatique À 2 SubstratsDocument7 paginiCinétique Enzymatique À 2 Substratsamina44Încă nu există evaluări
- Cinetique 2Document22 paginiCinetique 2BENARIMA AbdelhakimÎncă nu există evaluări
- TD+Corr Série 5Document6 paginiTD+Corr Série 5ismailmoudziri7Încă nu există evaluări
- Phot Cata Chapitre IDocument11 paginiPhot Cata Chapitre IHk EhÎncă nu există evaluări
- Fiche TD Enzymologie Réactions Enzymatiques - CopieDocument2 paginiFiche TD Enzymologie Réactions Enzymatiques - Copiebenali anfalÎncă nu există evaluări
- BCPST1 chpII-cinétiq 2022Document10 paginiBCPST1 chpII-cinétiq 2022Fannes DesignÎncă nu există evaluări
- But Du TPDocument12 paginiBut Du TPGüzęl Bârãæ100% (1)
- Cours La Cinétique ChimiqueDocument3 paginiCours La Cinétique ChimiqueBadre-ezzamen KaddourÎncă nu există evaluări
- 7 Mecanismes Bibi PDFDocument9 pagini7 Mecanismes Bibi PDFOussama IdrissÎncă nu există evaluări
- Cinétique EnzymatiqueDocument4 paginiCinétique EnzymatiqueAÎncă nu există evaluări
- DS04 CDocument10 paginiDS04 CbaatoutmbÎncă nu există evaluări
- NigriDocument4 paginiNigriأمير عطافيÎncă nu există evaluări
- Bio 322 BDocument4 paginiBio 322 BMourad DjeerafÎncă nu există evaluări
- TD Calcul Des Réacteurs Chimiques Master Matériaux FPNDocument37 paginiTD Calcul Des Réacteurs Chimiques Master Matériaux FPNHã Nã100% (2)
- Controle de Rattrapage de Cinétique 2019 2020 Avec CorrigéDocument7 paginiControle de Rattrapage de Cinétique 2019 2020 Avec CorrigéMy ProofÎncă nu există evaluări
- Cinetique Enz.a 2 Sub1Document14 paginiCinetique Enz.a 2 Sub1Mira45Încă nu există evaluări
- Cours 3 Cinétique Khenifi-ConvertiDocument6 paginiCours 3 Cinétique Khenifi-ConvertiÑar ĎjesÎncă nu există evaluări
- 1 Concepts CatalyseDocument15 pagini1 Concepts CatalyseAbdullah MohamedÎncă nu există evaluări
- Cinetique Chimique PDFDocument21 paginiCinetique Chimique PDFياسين السعدانيÎncă nu există evaluări
- Chapitre 2 La Cinétique EnzymatiqueDocument15 paginiChapitre 2 La Cinétique EnzymatiqueAbdouli RoukayaÎncă nu există evaluări
- Polycopié Cinétique Chimique L2 L3 GP en PDF Edit Par Dr. NouairiDocument83 paginiPolycopié Cinétique Chimique L2 L3 GP en PDF Edit Par Dr. NouairiOumateÎncă nu există evaluări
- Cinétique S5 2016Document122 paginiCinétique S5 2016Mohamed EL FAGHLOUMIÎncă nu există evaluări
- CH5 CinetiqueDocument17 paginiCH5 CinetiqueMétuschélah Lagouth100% (2)
- GCH 2006 Calcul Des Reacteurs Chimiques Cours PDFDocument16 paginiGCH 2006 Calcul Des Reacteurs Chimiques Cours PDFsettou bouthGpÎncă nu există evaluări
- 1S-T1.2Modélisation Et Suivi D'une Transformation Chimique Et Titrage ColorimétriqueDocument4 pagini1S-T1.2Modélisation Et Suivi D'une Transformation Chimique Et Titrage ColorimétriqueAlejandra Lorite AbadÎncă nu există evaluări
- TD EnzymologieDocument5 paginiTD EnzymologieAntadiop9Încă nu există evaluări
- TD 2 D'enzymologieDocument3 paginiTD 2 D'enzymologieabdel krim100% (2)
- td6 Cinetique ChimiqueDocument2 paginitd6 Cinetique ChimiqueAdri LebÎncă nu există evaluări
- TD Meca Reactionnelscorrige PDFDocument13 paginiTD Meca Reactionnelscorrige PDFAyyoub Dahbi100% (1)
- Cours - Chimie - RESUME CINETIQUE CHIMIQUE - Bac Sciences Exp (2015-2016) MR Daghsni SahbiDocument3 paginiCours - Chimie - RESUME CINETIQUE CHIMIQUE - Bac Sciences Exp (2015-2016) MR Daghsni SahbiKais Ben Aicha100% (2)
- Cinitique EnzymatiqueDocument11 paginiCinitique EnzymatiqueAkiliss VitchiÎncă nu există evaluări
- Fichier Produit 2658Document4 paginiFichier Produit 2658FlorinaÎncă nu există evaluări