S-ar putea să vă placă și
- Semana 1. Generalidades Bioenergética y TermodinámicaDocument30 paginiSemana 1. Generalidades Bioenergética y TermodinámicaMaria Paula Torres GarciaÎncă nu există evaluări
- Generalidades Bioenergética y TermodinámicaDocument37 paginiGeneralidades Bioenergética y TermodinámicaIván Gómez OcampoÎncă nu există evaluări
- 0tema 2 BioenergeticaDocument12 pagini0tema 2 BioenergeticaGema SantacreuÎncă nu există evaluări
- U1 ABP Bioenergética Gil Becerril ReyesDocument13 paginiU1 ABP Bioenergética Gil Becerril ReyesEdgar BecerrilÎncă nu există evaluări
- Clase IDocument35 paginiClase IdanÎncă nu există evaluări
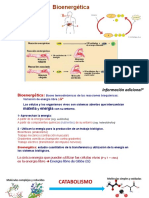
- BioenergéticaDocument84 paginiBioenergéticaLuisÎncă nu există evaluări
- Principios de BioenergeticaDocument15 paginiPrincipios de BioenergeticaRuth Yeimi Ochasara FrancoÎncă nu există evaluări
- 3clase Teórica Bioenergética 2023Document24 pagini3clase Teórica Bioenergética 2023Jos GmÎncă nu există evaluări
- Tema 8 - BioenergéticaDocument11 paginiTema 8 - BioenergéticaLara MaríaÎncă nu există evaluări
- Principios de BioenergeticaDocument27 paginiPrincipios de BioenergeticaRuth Yeimi Ochasara FrancoÎncă nu există evaluări
- EJERCICIOS DE ENTALPIA II-A Grupo 5Document9 paginiEJERCICIOS DE ENTALPIA II-A Grupo 5Yosef Camposano RodriguezÎncă nu există evaluări
- Guia 3-Energetica de La Vida 2018 ResueltaDocument5 paginiGuia 3-Energetica de La Vida 2018 ResueltaRocioÎncă nu există evaluări
- Termodinamica y BioenergéticaDocument52 paginiTermodinamica y BioenergéticaMartin BartolotteÎncă nu există evaluări
- Energia Libre GibbsDocument17 paginiEnergia Libre GibbsjuanvhoccÎncă nu există evaluări
- TermodinámicaDocument16 paginiTermodinámicaJonathan LealÎncă nu există evaluări
- T10 BioenergéticaDocument69 paginiT10 BioenergéticaAniri MaciasÎncă nu există evaluări
- "Bioenergética:": Libera ExotérmicaDocument21 pagini"Bioenergética:": Libera ExotérmicaMARÍA BARROSÎncă nu există evaluări
- Pc0 - Termodinámica (ATP y OB)Document55 paginiPc0 - Termodinámica (ATP y OB)Bathory LunaÎncă nu există evaluări
- Termodinamica y MetabolismoDocument16 paginiTermodinamica y Metabolismoequipo xochiÎncă nu există evaluări
- UNIDAD I - Energia Libre Gibbs-Equilibrio y Espontaneidad-Gibbs-2020Document44 paginiUNIDAD I - Energia Libre Gibbs-Equilibrio y Espontaneidad-Gibbs-2020Geronimo Israel MejiaÎncă nu există evaluări
- BioenergeticaDocument4 paginiBioenergeticaDANIELA ANDREA VARGAS ORTEGAÎncă nu există evaluări
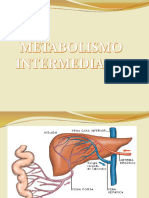
- Metabolismo Intermedio...Document18 paginiMetabolismo Intermedio...Dalia LucanaÎncă nu există evaluări
- Termodinamica MetabolismoDocument16 paginiTermodinamica MetabolismoAngel Sanchez FregosoÎncă nu există evaluări
- Capitulo 12 Bioenerg MED 15Document7 paginiCapitulo 12 Bioenerg MED 15TheBenjaxÎncă nu există evaluări
- Tema 6. Bioenergética y TermodinámicaDocument55 paginiTema 6. Bioenergética y TermodinámicaDarwin ValverdeÎncă nu există evaluări
- CLASE 4 de 2024-0Document42 paginiCLASE 4 de 2024-0fer.hidalvÎncă nu există evaluări
- Bioenergetica - Glucolisis - 2021 - PresentaciónDocument43 paginiBioenergetica - Glucolisis - 2021 - Presentaciónjulicatu90Încă nu există evaluări
- Termodinamica y Bioenergetica 1Document54 paginiTermodinamica y Bioenergetica 1cesar camberaÎncă nu există evaluări
- UNIDAD I-SESIONES 4Y5-Energia Libre Gibbs-Equilibrio y Espontaneidad-Gibbs-2019Document44 paginiUNIDAD I-SESIONES 4Y5-Energia Libre Gibbs-Equilibrio y Espontaneidad-Gibbs-2019Alma GarciaÎncă nu există evaluări
- Bioquimica Compuestos FosforiladosDocument5 paginiBioquimica Compuestos FosforiladosjuergenfrankÎncă nu există evaluări
- Taller de Repaso No.1 BioenergéticaDocument3 paginiTaller de Repaso No.1 BioenergéticaJuancho IbarraÎncă nu există evaluări
- BioenergeticaDocument43 paginiBioenergeticaChris Guzman100% (1)
- Termodinamica y Equilibrio QuimicoDocument31 paginiTermodinamica y Equilibrio QuimicoFranciscoÎncă nu există evaluări
- Energia Libre de Gibbs y Equilibrio Quimico2Document35 paginiEnergia Libre de Gibbs y Equilibrio Quimico2Elvis Jhordan Abad MirandaÎncă nu există evaluări
- Energía Libre de Gibbs y Equilibrio QuímicoDocument35 paginiEnergía Libre de Gibbs y Equilibrio QuímicotrigresvlaqsÎncă nu există evaluări
- Para Determinar Si Un Proceso Es o No EspontáneoDocument2 paginiPara Determinar Si Un Proceso Es o No EspontáneoEmmaColladoArredondoÎncă nu există evaluări
- Teoría FQ 07 Mat Set.23 2023-IIDocument18 paginiTeoría FQ 07 Mat Set.23 2023-IIsofia ines huaripata ponceÎncă nu există evaluări
- BIOENERGETICADocument47 paginiBIOENERGETICASofia Montiel Ochoa100% (1)
- BioenergeticaDocument25 paginiBioenergeticaadolfo moralesÎncă nu există evaluări
- Taller 2 Fisicoquímica 1Document17 paginiTaller 2 Fisicoquímica 1alf_1987Încă nu există evaluări
- Bioenergetica para Ing Biomed - 2019 PDFDocument82 paginiBioenergetica para Ing Biomed - 2019 PDFAlexsandraJ.CorderoDonaireÎncă nu există evaluări
- Problemas de TERMODINÁMICA y BIOENERGÉTICA 2024Document2 paginiProblemas de TERMODINÁMICA y BIOENERGÉTICA 2024Yanina TorresÎncă nu există evaluări
- Bioenergética 2020Document38 paginiBioenergética 2020Ángel Marcelo BravoÎncă nu există evaluări
- 2º Parcial BioquímicaDocument20 pagini2º Parcial BioquímicaFernanda Jacobo RodriguezÎncă nu există evaluări
- Resumen de TermoquímicaDocument11 paginiResumen de TermoquímicatotithhshshsgÎncă nu există evaluări
- PAU TermodinamicaDocument91 paginiPAU TermodinamicaPruden HidalgoÎncă nu există evaluări
- Sol Problemas Tema 4Document10 paginiSol Problemas Tema 4hugopocia23Încă nu există evaluări
- Transferencia de Calor en Las Reacciones QuímicasDocument16 paginiTransferencia de Calor en Las Reacciones QuímicasAlejandro Concha SalazarÎncă nu există evaluări
- BioenergéticaDocument14 paginiBioenergéticajoel panayÎncă nu există evaluări
- Termoquímica ApuntesDocument17 paginiTermoquímica ApuntesMarta RubioÎncă nu există evaluări
- BioenergeticaDocument50 paginiBioenergeticaRosMarie Tamayo67% (3)
- (Química 2º Bachillerato) Ejercicios + Soluciones: Energía de Las Reacciones Químicas. EspontaneidadDocument21 pagini(Química 2º Bachillerato) Ejercicios + Soluciones: Energía de Las Reacciones Químicas. EspontaneidadÁlvaro S.100% (5)
- Taller No.2 BioenergéticaDocument3 paginiTaller No.2 BioenergéticaEliana TrujilloÎncă nu există evaluări
- Segundo Pricipio de La TermodinamicaDocument14 paginiSegundo Pricipio de La TermodinamicaEbony Velasco LauraÎncă nu există evaluări
- Termodinamica - Segundo PrincipioDocument37 paginiTermodinamica - Segundo PrincipioNahuelÎncă nu există evaluări
- S11.s1 - Material - TermoquímicaDocument23 paginiS11.s1 - Material - TermoquímicaOlger Aragon BerlangaÎncă nu există evaluări
- Solución Problemas TermoquímicaDocument5 paginiSolución Problemas TermoquímicaInés D-PhantomÎncă nu există evaluări
- TermoquimicaDocument21 paginiTermoquimicajose serranoÎncă nu există evaluări
- Bioenergética PDFDocument6 paginiBioenergética PDFalexmaras95100% (1)
- Ingeniería química. Soluciones a los problemas del tomo IDe la EverandIngeniería química. Soluciones a los problemas del tomo IÎncă nu există evaluări
- Actividades Plan de Intervenciones Colectivas 2022Document9 paginiActividades Plan de Intervenciones Colectivas 2022Jesús BautistaÎncă nu există evaluări
- La Doctrina Del Consentimiento Informado en El Ordenamiento Jurídico Norteamericano PDFDocument22 paginiLa Doctrina Del Consentimiento Informado en El Ordenamiento Jurídico Norteamericano PDFJesús BautistaÎncă nu există evaluări
- Panel Viral RespiratorioDocument2 paginiPanel Viral RespiratorioJesús BautistaÎncă nu există evaluări
- Panel Viral RespiratorioDocument2 paginiPanel Viral RespiratorioJesús BautistaÎncă nu există evaluări
- Ad ADocument10 paginiAd AJesús BautistaÎncă nu există evaluări
- Ani MalesDocument56 paginiAni MalesJesús BautistaÎncă nu există evaluări
- La Célula y El MicroscopioDocument8 paginiLa Célula y El MicroscopiojulioebgÎncă nu există evaluări
- Desarrolle Una Conceptualización de MetabolismoDocument13 paginiDesarrolle Una Conceptualización de MetabolismoJohan MoreraÎncă nu există evaluări
- Capítulo 11 - Bioenergética - La Función Del ATPDocument6 paginiCapítulo 11 - Bioenergética - La Función Del ATPYissette Bautista CarrilloÎncă nu există evaluări
- Diversidad CatabolicaDocument9 paginiDiversidad CatabolicaAngg StumpÎncă nu există evaluări
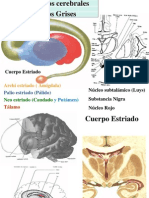
- BIOLOGIADocument57 paginiBIOLOGIAsantiagoÎncă nu există evaluări
- Biología IDocument9 paginiBiología IYasmin BarriosÎncă nu există evaluări
- Articulo TermodinamicaDocument15 paginiArticulo Termodinamicanathalycruz100% (1)
- Bombeo de Protones en Membrana Celular de Levaduras y Sus Procesos InhibitoriosDocument7 paginiBombeo de Protones en Membrana Celular de Levaduras y Sus Procesos InhibitoriosGabriela CoyÎncă nu există evaluări
- FOTOSÍNTESIS Exposición DocumentoDocument3 paginiFOTOSÍNTESIS Exposición DocumentoMaría J. FigueroaÎncă nu există evaluări
- Practicas Plantas 2 Oscar y PalomaDocument307 paginiPracticas Plantas 2 Oscar y PalomaCristian GallegoÎncă nu există evaluări
- Monitoreo y Control de BiofoulingDocument19 paginiMonitoreo y Control de Biofouling@sssÎncă nu există evaluări
- Evaluación Unidad 3 Ciencias de La Salud IDocument9 paginiEvaluación Unidad 3 Ciencias de La Salud IRutilioÎncă nu există evaluări
- Actividad 15 - BiologiaDocument2 paginiActividad 15 - BiologiaAna Natalia Pérez GaticaÎncă nu există evaluări
- Fenomenos de Reposicion Energetica de La ContraccionDocument3 paginiFenomenos de Reposicion Energetica de La ContraccionOriana Blanco100% (1)
- Evaluación SéptimoDocument4 paginiEvaluación SéptimoJuan orjuelaÎncă nu există evaluări
- Clase 3 Bioquimica Atp Itb Raynier PDFDocument41 paginiClase 3 Bioquimica Atp Itb Raynier PDFDennis MartinezÎncă nu există evaluări
- Conservacion de La Trucha PDFDocument137 paginiConservacion de La Trucha PDFraquelÎncă nu există evaluări
- CofactoresDocument17 paginiCofactoresDenisse MadridÎncă nu există evaluări
- Silabo 2023 MHDocument7 paginiSilabo 2023 MHJEAN FRANKO PINTO ASENCIOÎncă nu există evaluări
- Problemas 01 02Document43 paginiProblemas 01 02Beniamino De GiustiÎncă nu există evaluări
- EVALUACIÓN SÉPTIMO BIOLOGÍA - QUÍMICA 1-2-3 - GUÍA, para II Semestre2020Document2 paginiEVALUACIÓN SÉPTIMO BIOLOGÍA - QUÍMICA 1-2-3 - GUÍA, para II Semestre2020Alexis OrtegaÎncă nu există evaluări
- Capitulo 5 - BioquímicaDocument93 paginiCapitulo 5 - BioquímicaDerrick hardy Sandoval hurtadoÎncă nu există evaluări
- Solx Adm Unt 2023-I - Area DDocument45 paginiSolx Adm Unt 2023-I - Area DJUNIOR SANTIAGO RUBIO POMAÎncă nu există evaluări
- ApuntesDocument30 paginiApuntesLucas GarcíaÎncă nu există evaluări
- GLUCÓLISISDocument21 paginiGLUCÓLISISAngy PalmaÎncă nu există evaluări
- Plan de Tesis - Consumo de Suplementos, Uso de Anabólicos y TDCDocument58 paginiPlan de Tesis - Consumo de Suplementos, Uso de Anabólicos y TDCAlexander YanaricoÎncă nu există evaluări
- 3 Obtencion Industrial Del WhiskyDocument38 pagini3 Obtencion Industrial Del Whiskypaez lamaÎncă nu există evaluări
- Paradigmas Del EntrenamientoDocument11 paginiParadigmas Del EntrenamientoCarlos GarcíaÎncă nu există evaluări
- Acidemia PiruvicaDocument11 paginiAcidemia PiruvicafernandaÎncă nu există evaluări
- Cadena Respiratoria y Fosforilación OxidativaDocument26 paginiCadena Respiratoria y Fosforilación OxidativaDavid NavarroÎncă nu există evaluări