S-ar putea să vă placă și
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- NOVAGEN Prepare Sample PAGE SDS PDFDocument3 paginiNOVAGEN Prepare Sample PAGE SDS PDFJan Elysia GangeÎncă nu există evaluări
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (587)
- Higher Algebra - Hall & KnightDocument593 paginiHigher Algebra - Hall & KnightRam Gollamudi100% (2)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (894)
- I. Rules of Statutory Construction: Literal Interpretation/plain Meaning RuleDocument5 paginiI. Rules of Statutory Construction: Literal Interpretation/plain Meaning RuleJan Elysia GangeÎncă nu există evaluări
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Higher Algebra - Hall & KnightDocument593 paginiHigher Algebra - Hall & KnightRam Gollamudi100% (2)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- The Evolution of The Revolution: Week WeekDocument2 paginiThe Evolution of The Revolution: Week Weekrosa acostaÎncă nu există evaluări
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- Biotechnology and Its ApplicationsDocument12 paginiBiotechnology and Its ApplicationsHemant KumarÎncă nu există evaluări
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- Cell Cycle StagesDocument16 paginiCell Cycle StagesSean0% (1)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- The Structure and Functions of the NucleusDocument19 paginiThe Structure and Functions of the NucleusYuu Ayu'k LifestarÎncă nu există evaluări
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- 12 Chap 2 (Biological Molecules) F.SC 1st Year Biology Helping Notes PDFDocument10 pagini12 Chap 2 (Biological Molecules) F.SC 1st Year Biology Helping Notes PDFZainab fatimaÎncă nu există evaluări
- Biology Cornell Notes HWDocument3 paginiBiology Cornell Notes HWapi-330055989Încă nu există evaluări
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Car-T Cell 2 (7757)Document8 paginiCar-T Cell 2 (7757)Saradha PellatiÎncă nu există evaluări
- Bio Lab QuestionsDocument1 paginăBio Lab Questionsfuriex100% (2)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Test 1 BotnyDocument6 paginiTest 1 BotnyaligrehanakÎncă nu există evaluări
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- Blue WhiteDocument2 paginiBlue WhiteShyamala RamalingamÎncă nu există evaluări
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2219)
- 4 RazzaqueDocument10 pagini4 RazzaquesanÎncă nu există evaluări
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- G26interno BrochureDocument6 paginiG26interno BrochurecyberwmxÎncă nu există evaluări
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Scmra: A Robust Deep Learning Method To Annotate Scrna-Seq Data With Multiple Reference DatasetsDocument15 paginiScmra: A Robust Deep Learning Method To Annotate Scrna-Seq Data With Multiple Reference DatasetsNavid ArefanÎncă nu există evaluări
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (265)
- 1Document141 pagini1DIXLVC XXIÎncă nu există evaluări
- Cloud Applications: Protein Structure Predication and Gene Expression Data AnalysisDocument10 paginiCloud Applications: Protein Structure Predication and Gene Expression Data AnalysisAyushmaan BadkulÎncă nu există evaluări
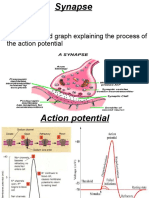
- Synapse Structure and Function ExplainedDocument11 paginiSynapse Structure and Function ExplainedMuhammad AbdullahÎncă nu există evaluări
- RyanodineReceptor1 RelatedMyop PDFDocument15 paginiRyanodineReceptor1 RelatedMyop PDFkay_mike_83Încă nu există evaluări
- Nervous System: Team MembersDocument8 paginiNervous System: Team MembersnehaÎncă nu există evaluări
- Transport MechanismsDocument2 paginiTransport MechanismsErica GarciaÎncă nu există evaluări
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- InFusion Cloning User ManualDocument15 paginiInFusion Cloning User ManualHannaÎncă nu există evaluări
- Drugs in The Pipeline For HBVDocument21 paginiDrugs in The Pipeline For HBVMarius StancuÎncă nu există evaluări
- Biochemistry (Chapter 1: A Brief History of Biochemistry) : March 2016Document31 paginiBiochemistry (Chapter 1: A Brief History of Biochemistry) : March 2016Biochem M. JulyÎncă nu există evaluări
- Introduction To Databases - NCBI, PDB and UniprotDocument5 paginiIntroduction To Databases - NCBI, PDB and UniprotMehak MattooÎncă nu există evaluări
- BIO 205 Chapter 8 PowerpointDocument16 paginiBIO 205 Chapter 8 PowerpointDrPearcyÎncă nu există evaluări
- Enzyme InhibitionDocument7 paginiEnzyme InhibitionJane Docdoc100% (1)
- h2 Mathematics Summarized Formulae3Document13 paginih2 Mathematics Summarized Formulae3Jin Yang ChewÎncă nu există evaluări
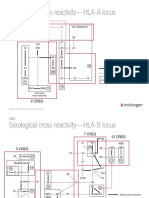
- Serological cross-reactivity-HLA-A Locus: 1 CregDocument5 paginiSerological cross-reactivity-HLA-A Locus: 1 CregGeorgia MarcussoÎncă nu există evaluări
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (119)
- History of DNA and the Central DogmaDocument1 paginăHistory of DNA and the Central Dogmaemarie8panerioÎncă nu există evaluări
- J. Biol. Chem.-1988-Meister-17205-8Document4 paginiJ. Biol. Chem.-1988-Meister-17205-8Pepé DubÎncă nu există evaluări
- Peptide HandbookDocument26 paginiPeptide HandbookKim HiềnÎncă nu există evaluări
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)