S-ar putea să vă placă și
- Immunology - Chapter 19 VaccinesDocument21 paginiImmunology - Chapter 19 VaccinesJJHHJHi100% (1)
- Lab 18: Immunology 18.1 The ELISA: Activity 1: Using An ELISA To Diagnose Lupus To BeginDocument6 paginiLab 18: Immunology 18.1 The ELISA: Activity 1: Using An ELISA To Diagnose Lupus To BeginValerie OkakpuÎncă nu există evaluări
- Pathophysiology - Step by Step G - Albert Nolan PHDDocument171 paginiPathophysiology - Step by Step G - Albert Nolan PHDΑΘΑΝΑΣΙΟΣ ΚΟΥΤΟΥΚΤΣΗΣ71% (7)
- Cellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsDe la EverandCellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsCharles G. CochraneÎncă nu există evaluări
- InterferonDocument6 paginiInterferonMonika SharmaÎncă nu există evaluări
- Core Topic 4 - The Different Types of Vaccines Used and Their Composition - March 2009Document27 paginiCore Topic 4 - The Different Types of Vaccines Used and Their Composition - March 2009Pranav NakhateÎncă nu există evaluări
- Bacterial Genetics: DR - Mohiruddin SK Senior Resident Dept of MicrobiologyDocument49 paginiBacterial Genetics: DR - Mohiruddin SK Senior Resident Dept of MicrobiologyAbcdefg HijklÎncă nu există evaluări
- Production of Humulin by Recombinant EDocument9 paginiProduction of Humulin by Recombinant EbsreegiriprasadÎncă nu există evaluări
- 1.0 ADR Reporting and Management PDFDocument35 pagini1.0 ADR Reporting and Management PDFDr. Raghavendra Kumar Gunda100% (1)
- 11.ethics CommiteeDocument18 pagini11.ethics CommiteeDRx Sonali TareiÎncă nu există evaluări
- FermentationDocument32 paginiFermentationJsquareÎncă nu există evaluări
- Immune System To Agent InfectionDocument70 paginiImmune System To Agent Infectionliza yunitaÎncă nu există evaluări
- Antibody and AntigenDocument22 paginiAntibody and Antigenwildan pratamaÎncă nu există evaluări
- 1.4.2 Making VaccinesDocument21 pagini1.4.2 Making VaccinesVikas Viki0% (1)
- Immunology: Chapter 1: An Overview of The Immune SystemDocument52 paginiImmunology: Chapter 1: An Overview of The Immune SystemMekuriya BeregaÎncă nu există evaluări
- MutationDocument33 paginiMutationarslan akramÎncă nu există evaluări
- Production of PenicillinDocument16 paginiProduction of PenicillinMeenal KhannaÎncă nu există evaluări
- Lab 6 Aseptic TechniqueDocument22 paginiLab 6 Aseptic TechniqueKrsna NaveraÎncă nu există evaluări
- Recent Trends in Viral VaccinesDocument64 paginiRecent Trends in Viral VaccinesAkula Sai PrasannaÎncă nu există evaluări
- Clinical TrialDocument26 paginiClinical TrialFatonyÎncă nu există evaluări
- Antibodies CHPT 4Document27 paginiAntibodies CHPT 4aswanÎncă nu există evaluări
- Bi 341 Chapter 7 Bacterial GeneticsDocument39 paginiBi 341 Chapter 7 Bacterial GeneticsMATHIXÎncă nu există evaluări
- Bacterial Genetics & ReproductionDocument18 paginiBacterial Genetics & ReproductionAsimÎncă nu există evaluări
- Vaccine Development, Testing, and RegulationDocument12 paginiVaccine Development, Testing, and RegulationP Bijoya SinghaÎncă nu există evaluări
- Blotting Techniques: The Muscular SystemDocument33 paginiBlotting Techniques: The Muscular SystemPrasanna BabuÎncă nu există evaluări
- Ich - GCP: Lets Discuss It BrieflyDocument9 paginiIch - GCP: Lets Discuss It Brieflyaks_pÎncă nu există evaluări
- Next Generation AntibodiesDocument30 paginiNext Generation AntibodiesMeitei IngobaÎncă nu există evaluări
- Hypersensitivity 1Document30 paginiHypersensitivity 1Oral Biology100% (1)
- Protocol and Case Record FormDocument29 paginiProtocol and Case Record Formsimple_anandÎncă nu există evaluări
- Specific Host Defenses: The Immune ResponseDocument54 paginiSpecific Host Defenses: The Immune Responseadyaly44Încă nu există evaluări
- Biochemical Engineering: Dr. Syed Nadir HussainDocument46 paginiBiochemical Engineering: Dr. Syed Nadir Hussainumairgul841Încă nu există evaluări
- Who, What, When, Where, How, and Why: The Ingredients in The Recipe For A Successful Methods SectionDocument5 paginiWho, What, When, Where, How, and Why: The Ingredients in The Recipe For A Successful Methods SectionMarilo Rus GuerreroÎncă nu există evaluări
- Vaccine: - Chhabi Acharya Binod Rasaili Rupesh K.C Hope Int'l CollegeDocument21 paginiVaccine: - Chhabi Acharya Binod Rasaili Rupesh K.C Hope Int'l CollegeScott Domes100% (1)
- AdenovirusDocument18 paginiAdenovirusNovika Ayuni RambeÎncă nu există evaluări
- Microbial Genetics TortoraDocument3 paginiMicrobial Genetics TortoraMiss DÎncă nu există evaluări
- Adaptive ImmunityDocument34 paginiAdaptive ImmunityMohamed Mido100% (1)
- Edible VaccinesDocument21 paginiEdible VaccinesPritish SareenÎncă nu există evaluări
- Immunoglobulins ClassDocument16 paginiImmunoglobulins ClassKoushali BanerjeeÎncă nu există evaluări
- Vaccine Production (L1F17PHMD0174)Document22 paginiVaccine Production (L1F17PHMD0174)Faizah Khalid100% (1)
- TransductionDocument14 paginiTransductionArchana RakibeÎncă nu există evaluări
- Introduction To Clinical ResearchDocument19 paginiIntroduction To Clinical ResearchVignesh GaneshÎncă nu există evaluări
- Application of rDNA in Animal Cell Culture (Animal Biotech)Document64 paginiApplication of rDNA in Animal Cell Culture (Animal Biotech)jithinnxÎncă nu există evaluări
- Presented by - Vasudha Singh Moderated by - Dr. Niti SinghalDocument42 paginiPresented by - Vasudha Singh Moderated by - Dr. Niti SinghalAvi VermaÎncă nu există evaluări
- Good Clinical Practice WorkshopDocument24 paginiGood Clinical Practice WorkshopDian Hapsari100% (1)
- Vaccine & Principle of ImmunizationDocument23 paginiVaccine & Principle of ImmunizationSindhu Babu100% (1)
- 4.10.17final Clinical Trials Talk.4.10.2017 - 300526 - 284 - 30426 - v1Document67 pagini4.10.17final Clinical Trials Talk.4.10.2017 - 300526 - 284 - 30426 - v1Mohammed HammedÎncă nu există evaluări
- Cell CultureDocument33 paginiCell CultureSai SridharÎncă nu există evaluări
- Molecular Biology Course Genetic VectorsDocument20 paginiMolecular Biology Course Genetic VectorsDr. Adnan Al-KanaanÎncă nu există evaluări
- Bacterial VaccineDocument44 paginiBacterial VaccineShrenik LuniyaÎncă nu există evaluări
- Industrial Importance of MicrobesDocument23 paginiIndustrial Importance of MicrobesDiah AyuningrumÎncă nu există evaluări
- New Developments in Chromogenic and Fluorogenic Culture MediaDocument14 paginiNew Developments in Chromogenic and Fluorogenic Culture MediaPaulina LizzethÎncă nu există evaluări
- Vaccines Science PharmaDocument30 paginiVaccines Science PharmaGanesh V GaonkarÎncă nu există evaluări
- Antibody EngineeringDocument14 paginiAntibody EngineeringJholina Aure ReyesÎncă nu există evaluări
- Bacterial Genetics: Calvin Bisong EbaiDocument58 paginiBacterial Genetics: Calvin Bisong EbaiMBAH LOIS LA GRACEÎncă nu există evaluări
- Difference Between ICH-GCP and Indian GCP Guidelines PGIDocument6 paginiDifference Between ICH-GCP and Indian GCP Guidelines PGINeha DungÎncă nu există evaluări
- Antigens & Antigen Presentation: Prof Anura WeerasingheDocument21 paginiAntigens & Antigen Presentation: Prof Anura Weerasinghesiva100% (1)
- Blotting TechniquesDocument3 paginiBlotting TechniquesabidaiqbalansariÎncă nu există evaluări
- Alternative Culture Media For Bacterial Growth Using Different Formulation of Protein SourcesDocument4 paginiAlternative Culture Media For Bacterial Growth Using Different Formulation of Protein SourcesMasitha Dwi AmiraÎncă nu există evaluări
- Gene Transfer MechanismDocument21 paginiGene Transfer MechanismNik SharmaÎncă nu există evaluări
- Molecular Diagnostics in Hematological OncologyDe la EverandMolecular Diagnostics in Hematological OncologyÎncă nu există evaluări
- Monoclonal Antibodies Against Bacteria: Volume IIIDe la EverandMonoclonal Antibodies Against Bacteria: Volume IIIAlberto J. L. MacarioÎncă nu există evaluări
- Rembe 1Document8 paginiRembe 1Steve ColbertÎncă nu există evaluări
- Histology Points of Indentifications 1st Year-1Document3 paginiHistology Points of Indentifications 1st Year-1SikandarÎncă nu există evaluări
- Anatomy and PhysiologyDocument2 paginiAnatomy and PhysiologyKathzkaMaeAgcaoiliÎncă nu există evaluări
- Medical Terminology For Medical Transcription TraineesDocument74 paginiMedical Terminology For Medical Transcription TraineesRangothri Sreenivasa SubramanyamÎncă nu există evaluări
- 1.1 AP 1 Lab Language of AnatomyDocument41 pagini1.1 AP 1 Lab Language of AnatomyTinxi TinÎncă nu există evaluări
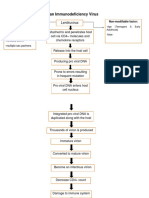
- Human Immunodeficiency Virus: LentiruvirusDocument5 paginiHuman Immunodeficiency Virus: Lentiruvirusjoyrena ochondraÎncă nu există evaluări
- Parasites Immunity and PathologyDocument389 paginiParasites Immunity and PathologyCarlos Bell Castro Tamayo100% (1)
- Identify The Choice That Best Completes The Statement or Answers The QuestionDocument14 paginiIdentify The Choice That Best Completes The Statement or Answers The QuestionCamille BaybayÎncă nu există evaluări
- IMMUNITYDocument23 paginiIMMUNITYFlorence Lynn BaisacÎncă nu există evaluări
- Q2 GPA Diseases of WBCs Lymph Nodes Spleen and ThymusDocument7 paginiQ2 GPA Diseases of WBCs Lymph Nodes Spleen and ThymusAdrian CaballesÎncă nu există evaluări
- Lymphatic System Transes 1Document8 paginiLymphatic System Transes 1Sophia Margarette CawalingÎncă nu există evaluări
- ANATOMY AND PHYSIOLOGY-flashcardsDocument21 paginiANATOMY AND PHYSIOLOGY-flashcardsJohn Wayne PharrÎncă nu există evaluări
- Beginner's Guide To YOUR LYMPHATICS 10 Common Things Thatclog Your Lymph 10 Signs & Symptoms of Lymph CongestionDocument14 paginiBeginner's Guide To YOUR LYMPHATICS 10 Common Things Thatclog Your Lymph 10 Signs & Symptoms of Lymph CongestionElvaÎncă nu există evaluări
- Assessing The Cardiovascular and Lymphatic SystemsDocument9 paginiAssessing The Cardiovascular and Lymphatic SystemsYAMINIPRIYANÎncă nu există evaluări
- Limfoproliferari Cronice MaligneDocument18 paginiLimfoproliferari Cronice MaligneAlice MuscaÎncă nu există evaluări
- Emily Iker, MD: LymphedemaDocument37 paginiEmily Iker, MD: LymphedemaJustin HarperÎncă nu există evaluări
- Hodgkin's DiseaseDocument5 paginiHodgkin's DiseaseKim PaoÎncă nu există evaluări
- 5 Anatomy of The Spleen DiyaDocument25 pagini5 Anatomy of The Spleen DiyaBra KojoÎncă nu există evaluări
- Anatomy LE2 Samplex 2017BDocument7 paginiAnatomy LE2 Samplex 2017BHanako Sasaki AranillaÎncă nu există evaluări
- Blood Physiology 2022Document116 paginiBlood Physiology 2022Gurmessa FekaduÎncă nu există evaluări
- Clauneta MondlaneDocument7 paginiClauneta MondlaneMiguelOriellÎncă nu există evaluări
- Leukemias and Lymphomas Flow Chart ModifiedDocument5 paginiLeukemias and Lymphomas Flow Chart Modifiedlovelyc95Încă nu există evaluări
- Treatment of Hodgkin Lymphoma - Non Hodgkin Lymphoma Treatment in India: Omegahospitals, HyderabadDocument2 paginiTreatment of Hodgkin Lymphoma - Non Hodgkin Lymphoma Treatment in India: Omegahospitals, HyderabadRyan ThompsonÎncă nu există evaluări
- Case Study On LeukemiaDocument55 paginiCase Study On LeukemialicservernoidaÎncă nu există evaluări
- Lymphatics of ThoraxDocument29 paginiLymphatics of ThoraxconovastyleÎncă nu există evaluări
- Introductiontothe Immune SystemDocument25 paginiIntroductiontothe Immune SystemUyên HuỳnhÎncă nu există evaluări
- NF-KB and The Immune ResponseDocument23 paginiNF-KB and The Immune ResponseAndri Praja SatriaÎncă nu există evaluări
- B.sc. NSG - Sylabus KUHASDocument63 paginiB.sc. NSG - Sylabus KUHASrijumathewmscÎncă nu există evaluări
- 1021 Lecture Question Bank PDFDocument44 pagini1021 Lecture Question Bank PDFaskldhfdasjkÎncă nu există evaluări