S-ar putea să vă placă și
- Ţesuturile EpitelialeDocument5 paginiŢesuturile Epitelialesura1234Încă nu există evaluări
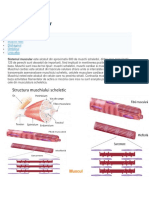
- 6.muscular 1-CorectatDocument13 pagini6.muscular 1-CorectatOana-Cristina CeacaÎncă nu există evaluări
- AnatomieDocument67 paginiAnatomieMarin GolbanÎncă nu există evaluări
- Muschii - GeneralitatiDocument13 paginiMuschii - GeneralitatiAna SanduÎncă nu există evaluări
- Anatomie Curs 3 Sau 4Document5 paginiAnatomie Curs 3 Sau 4Liviu GhilanÎncă nu există evaluări
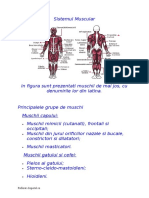
- Sistemul MuscularDocument30 paginiSistemul Muscularmidnightpoison1Încă nu există evaluări
- 01 CursDocument16 pagini01 CursmariaÎncă nu există evaluări
- Sistem MuschiDocument19 paginiSistem MuschiDan MassonÎncă nu există evaluări
- MușchiiDocument8 paginiMușchiiКарина Городецкая100% (1)
- Anatomia Si Fiziologia Sistemului MuscularDocument36 paginiAnatomia Si Fiziologia Sistemului MuscularGligan MihaiÎncă nu există evaluări
- MiologiaDocument107 paginiMiologiaamarieirazvan29Încă nu există evaluări
- Curs 9 Tesuturi MusculareDocument11 paginiCurs 9 Tesuturi MusculareAndra GarbeaÎncă nu există evaluări
- Miologie GeneralaDocument2 paginiMiologie GeneralaAnamaria HaneaÎncă nu există evaluări
- Organismul Uman Și SănătateaDocument16 paginiOrganismul Uman Și SănătateaNicoleta CobanÎncă nu există evaluări
- Muschi ScheleticiDocument45 paginiMuschi Scheleticizahariagheorghe100% (6)
- 5 - Tesutul MuscularDocument2 pagini5 - Tesutul MuscularDiana PetreaÎncă nu există evaluări
- Muşchiul Şi Contracţia MuscularăDocument9 paginiMuşchiul Şi Contracţia MuscularăIonut CojoÎncă nu există evaluări
- Miologie (Muschi, Tendon)Document38 paginiMiologie (Muschi, Tendon)Marina CarmenÎncă nu există evaluări
- Sistemul Muscular AnatomieDocument6 paginiSistemul Muscular Anatomiemihai bibiÎncă nu există evaluări
- MuschiulDocument11 paginiMuschiulviorelÎncă nu există evaluări
- Anatomie Toate CursurileDocument473 paginiAnatomie Toate CursurileGHIBERDICU IONUT FLORINÎncă nu există evaluări
- Miologie Curs IIIDocument41 paginiMiologie Curs IIIOpris AndreiÎncă nu există evaluări
- Referat Anatomie 3Document6 paginiReferat Anatomie 3Mihnea BucurÎncă nu există evaluări
- Tema7 Notiuni Fundamentale Privind Sistemul MuscularDocument9 paginiTema7 Notiuni Fundamentale Privind Sistemul MuscularOnofrei Oana ElenaÎncă nu există evaluări
- Tema 10Document8 paginiTema 10Denis CozlovÎncă nu există evaluări
- Fiziopatologia MuschiuluiDocument11 paginiFiziopatologia Muschiuluisorin1982Încă nu există evaluări
- TMK Curs ViiiDocument10 paginiTMK Curs ViiiviorelÎncă nu există evaluări
- Structura Muschilor.Document7 paginiStructura Muschilor.Yarik Botnar33% (3)
- Anatomie Curs 3Document4 paginiAnatomie Curs 3Costiuc LuminitaÎncă nu există evaluări
- TesuturiDocument25 paginiTesuturiDenisa AlexandraÎncă nu există evaluări
- Tesuturi MusculareDocument12 paginiTesuturi MusculareAlexandroae GabrielaÎncă nu există evaluări
- C 1 Sistemul NeuromuscularDocument67 paginiC 1 Sistemul NeuromuscularAlexandru MindruÎncă nu există evaluări
- AnatomieDocument4 paginiAnatomieAlexandrina DamianÎncă nu există evaluări
- 9.biomecanica - Ap. LocomotorDocument3 pagini9.biomecanica - Ap. LocomotorAndra BadescuÎncă nu există evaluări
- Tesutul Muscular Scheletic - BFK - 2022 PDFDocument45 paginiTesutul Muscular Scheletic - BFK - 2022 PDFLorena StroeÎncă nu există evaluări
- Prezentare MIOLOGIADocument60 paginiPrezentare MIOLOGIAAnca KynkÎncă nu există evaluări
- Articulatia Este Ansamblul Partilor Moi Prin Care Se Unesc Doua Sau Mai Multe Oase Vecine. ClasificareaDocument3 paginiArticulatia Este Ansamblul Partilor Moi Prin Care Se Unesc Doua Sau Mai Multe Oase Vecine. ClasificareaChiracuAndreeaÎncă nu există evaluări
- Ţesutul MuscularDocument3 paginiŢesutul MuscularGeorgiana TirnovanÎncă nu există evaluări
- Curs 3miologieDocument37 paginiCurs 3miologieVlaicu Tudor100% (1)
- Sistemul MuscularDocument18 paginiSistemul MuscularPasionat MasiniÎncă nu există evaluări
- AMG Anul I Anatomie 2Document3 paginiAMG Anul I Anatomie 2Beatrix Kudor100% (1)
- Tesut Muscular 26324Document111 paginiTesut Muscular 26324Galina BotnariÎncă nu există evaluări
- Curs 6 MiologiaDocument23 paginiCurs 6 MiologiaTrixie BeaÎncă nu există evaluări
- Biologie Celulara Si HistologieDocument78 paginiBiologie Celulara Si HistologieRaluca StefanescuÎncă nu există evaluări
- Tesutul MuscularDocument5 paginiTesutul MuscularIacob Paula-IzabelaÎncă nu există evaluări
- 1 ContractiaDocument10 pagini1 ContractiaAlina VoicuÎncă nu există evaluări
- Tesuturi MusculateDocument40 paginiTesuturi MusculateIoana NavaliciÎncă nu există evaluări
- Curs 7 ANATOMIEDocument4 paginiCurs 7 ANATOMIECezar PetreaÎncă nu există evaluări
- Școala Postliceală FDocument11 paginiȘcoala Postliceală FCristina GabrielaÎncă nu există evaluări
- Grupe Muschi PrincipaleDocument4 paginiGrupe Muschi Principaleandreea barosanÎncă nu există evaluări
- Test OsosDocument3 paginiTest OsosRaluca Moroșan100% (1)
- An III MusculatieDocument128 paginiAn III MusculatieRares Solcanu100% (1)
- Tesutul Muscular PDFDocument26 paginiTesutul Muscular PDFSpinu LiliaÎncă nu există evaluări
- Caracteristicile Generale Ale Ţesutului Muscular PDFDocument14 paginiCaracteristicile Generale Ale Ţesutului Muscular PDFsgaftonianuÎncă nu există evaluări
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 4 din 5 stele4/5 (8)
- Microcirculația sanguină: Următoarea generație de asistență socialăDe la EverandMicrocirculația sanguină: Următoarea generație de asistență socialăÎncă nu există evaluări
- Ai grija de viata, de tine, de ceilalti si de naturaDe la EverandAi grija de viata, de tine, de ceilalti si de naturaÎncă nu există evaluări
- Speranța nemuririi: Reflecții despre moarte și supraviețuireDe la EverandSperanța nemuririi: Reflecții despre moarte și supraviețuireÎncă nu există evaluări