S-ar putea să vă placă și
- Istoria de sub covor. Dezbateri istoriograficeDe la EverandIstoria de sub covor. Dezbateri istoriograficeEvaluare: 3.5 din 5 stele3.5/5 (3)
- Cursuri universitare de arheologie preistoricăDe la EverandCursuri universitare de arheologie preistoricăEvaluare: 5 din 5 stele5/5 (2)
- 096 Pontica 5 (1972) M.Document18 pagini096 Pontica 5 (1972) M.cosacmarianÎncă nu există evaluări
- De La Arhaic La Precambrian - Cariera de La Cosăuți: o Secțiune Geologică RevizuităDocument13 paginiDe La Arhaic La Precambrian - Cariera de La Cosăuți: o Secțiune Geologică RevizuităIgor NicoaraÎncă nu există evaluări
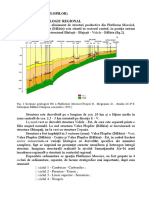
- Litologia Si Si Biostratigrafia Cretacicului Inferior Din Zona Muntilor BucegiDocument8 paginiLitologia Si Si Biostratigrafia Cretacicului Inferior Din Zona Muntilor Bucegidan manÎncă nu există evaluări
- Puiu Hasotti, Epoca Neolitica in Dobrogea PDFDocument315 paginiPuiu Hasotti, Epoca Neolitica in Dobrogea PDFAndreiIulianChirilă100% (3)
- Studiul Geologic Muntii Padurea Craiului Vol 3-1966Document240 paginiStudiul Geologic Muntii Padurea Craiului Vol 3-1966Denisa DobrescuÎncă nu există evaluări
- Vasile Chirica, Octavian Liviu Şovan, Civilisations Prehistoriques Et Protohistoriques de La Zone Du Prut MoyenDocument68 paginiVasile Chirica, Octavian Liviu Şovan, Civilisations Prehistoriques Et Protohistoriques de La Zone Du Prut MoyenCIMECÎncă nu există evaluări
- Ape Subterane PDFDocument20 paginiApe Subterane PDFAndreeaÎncă nu există evaluări
- HartageolDocument8 paginiHartageollidmary2001Încă nu există evaluări
- 03 Peuce SV Stiintele NaturiiDocument20 pagini03 Peuce SV Stiintele NaturiiAndreea TeleșpanÎncă nu există evaluări
- Distingerea de MoreneDocument2 paginiDistingerea de MoreneOvidiu VÎncă nu există evaluări
- Alcanii cu catena ramificată au numele format din numele catenei laterale notat ca prefix la numele celei mai lungi catene prezente în formulă. Poziţia catenei laterale seindică printr-un număr. Pentru aceasta, catena cea mai lungă se numerotează de la uncapăt la altul, cu numere arable, în acea direcţie în care catena laterală să ocupe poziţiacu numărul cel mai mic. Prezenţa mai multor catene laterale identice se indică prinDocument19 paginiAlcanii cu catena ramificată au numele format din numele catenei laterale notat ca prefix la numele celei mai lungi catene prezente în formulă. Poziţia catenei laterale seindică printr-un număr. Pentru aceasta, catena cea mai lungă se numerotează de la uncapăt la altul, cu numere arable, în acea direcţie în care catena laterală să ocupe poziţiacu numărul cel mai mic. Prezenţa mai multor catene laterale identice se indică prinStrack DanielÎncă nu există evaluări
- Orasul Gaesti - Cercetare Geografica Si GeologicaDocument116 paginiOrasul Gaesti - Cercetare Geografica Si GeologicapetrvasiÎncă nu există evaluări
- Studiu DambovicioaraDocument7 paginiStudiu DambovicioaraGrigorovici IonÎncă nu există evaluări
- RezumatDocument59 paginiRezumatAna Maria RusuÎncă nu există evaluări
- BalariaDocument12 paginiBalariaMaryllena MaryÎncă nu există evaluări
- Calcarul de RonaDocument4 paginiCalcarul de RonaAndreka AdrianÎncă nu există evaluări
- Vulcanismul NeogenDocument15 paginiVulcanismul NeogenCornel DitaÎncă nu există evaluări
- Statia Electrica Domnesti Caracterizare Geologica, Hidrogeologica Si ClimaticaDocument6 paginiStatia Electrica Domnesti Caracterizare Geologica, Hidrogeologica Si ClimaticaDutu MihaiÎncă nu există evaluări
- Studiul Geologic Al Dolomitelor Masive Hydaspiene Din Sinclinalul Haghimas (Panza Bucovinica)Document9 paginiStudiul Geologic Al Dolomitelor Masive Hydaspiene Din Sinclinalul Haghimas (Panza Bucovinica)leniucvasileÎncă nu există evaluări
- 07 Bucuresti Materiale de Istorie Si Muzeografie VII 1969Document315 pagini07 Bucuresti Materiale de Istorie Si Muzeografie VII 1969Biblioteca Targoviste100% (1)
- 8 Boroffka ApulumDocument24 pagini8 Boroffka ApulumBogdan BobinaÎncă nu există evaluări
- Orasul Gaesti - Cercetare Geografica Si GeologicaDocument116 paginiOrasul Gaesti - Cercetare Geografica Si Geologicagiugiuca1100% (1)
- Curs 6Document7 paginiCurs 6TEOFIL BÎRAÎncă nu există evaluări
- Cap.2 Geologia - Culoarul Bran-Rucar-Dragoslavele Studiu de Geografie Fizica Si Evaluarea PeisajuluiDocument8 paginiCap.2 Geologia - Culoarul Bran-Rucar-Dragoslavele Studiu de Geografie Fizica Si Evaluarea Peisajuluigeorgebala100% (2)
- 034 Cercetari Istorice Muzeul Istorie Moldovei Iasi 2015 - 038Document71 pagini034 Cercetari Istorice Muzeul Istorie Moldovei Iasi 2015 - 038teddy1967Încă nu există evaluări
- Geologia Romaniei, Vol I, II.2. Platforma SciticaDocument4 paginiGeologia Romaniei, Vol I, II.2. Platforma SciticaAnca SesermanÎncă nu există evaluări
- Cateva Materiale Din Neoliticul TimpuriuDocument10 paginiCateva Materiale Din Neoliticul Timpuriuffd jhkyÎncă nu există evaluări
- Geologie StructuralaDocument186 paginiGeologie StructuralazgributzÎncă nu există evaluări
- Geol Romaniei 2012 PDFDocument99 paginiGeol Romaniei 2012 PDFCesonia C DimitriaÎncă nu există evaluări
- RMM - MIA - 1977 - Anul XLVI - Nr2Document98 paginiRMM - MIA - 1977 - Anul XLVI - Nr2Nedelcu AndreiÎncă nu există evaluări
- Lubenescu Et Al., 1986 - Stratigrafie TRDocument6 paginiLubenescu Et Al., 1986 - Stratigrafie TRŞtefan VasileÎncă nu există evaluări
- Geologia Romaniei - Curs 04 - Platforma SciticaDocument17 paginiGeologia Romaniei - Curs 04 - Platforma SciticaAlexandru MiroiuÎncă nu există evaluări
- Dinamica Lacului Tei Intre Anii 1910-2005Document17 paginiDinamica Lacului Tei Intre Anii 1910-2005dots_rodi100% (1)
- Cercetarea Arheologica de SuprafataDocument6 paginiCercetarea Arheologica de SuprafataCUZIC VLADÎncă nu există evaluări
- Substratul GeologicDocument16 paginiSubstratul GeologicSofia Ciocan GraurÎncă nu există evaluări
- Industria Materialelor Osoase in Paleolitic, Pe Versantul CarpatilorDocument28 paginiIndustria Materialelor Osoase in Paleolitic, Pe Versantul Carpatilormarandan85Încă nu există evaluări
- Platforma Deltei Dunarii Prezentare GeologicaDocument28 paginiPlatforma Deltei Dunarii Prezentare GeologicaMarianaÎncă nu există evaluări
- Ape SubteraneDocument51 paginiApe SubteraneJusst X Mequerty100% (1)
- Geologia Romaniei - Curs 03 - Platforma MoldoveneascaDocument14 paginiGeologia Romaniei - Curs 03 - Platforma MoldoveneascaMarin Aostacioae100% (1)
- Studiul Asupra Unor Capcane Nestructurale Din Platform MoesicaDocument10 paginiStudiul Asupra Unor Capcane Nestructurale Din Platform MoesicaFloarea AlexandruÎncă nu există evaluări
- Geologia Carpatilor OrientaliDocument11 paginiGeologia Carpatilor Orientalidan man100% (2)
- Relief Carstic Geomorfologie 1970Document19 paginiRelief Carstic Geomorfologie 1970daniel193Încă nu există evaluări
- Patru-Stupariu Ileana Georgeta, Culoarul Transcarpatic Bran-Rucar-Dragoslavele - Studiu de Geografie Fizica, Cap. 2Document9 paginiPatru-Stupariu Ileana Georgeta, Culoarul Transcarpatic Bran-Rucar-Dragoslavele - Studiu de Geografie Fizica, Cap. 2georgebalaÎncă nu există evaluări
- Proiect 2Document13 paginiProiect 2Prisaca CristianÎncă nu există evaluări
- Inceputurile Geologiei in RomaniaDocument23 paginiInceputurile Geologiei in RomaniaSarbu AnamariaÎncă nu există evaluări
- 2 Sectiunea GeologieDocument25 pagini2 Sectiunea GeologiePaulina GrunwaldÎncă nu există evaluări
- Biocenozele Bentale de Pe Substratul Dur Din Sudul Litoralului RomanescDocument26 paginiBiocenozele Bentale de Pe Substratul Dur Din Sudul Litoralului RomanescObergefreiter Ciucardel prin mila lui DumnezeuÎncă nu există evaluări
- Geologia Dolomita Paraul CailorDocument8 paginiGeologia Dolomita Paraul CailorleniucvasileÎncă nu există evaluări
- Istoria Geologica A Merii Sarmatiene in Viziunea ActualaDocument6 paginiIstoria Geologica A Merii Sarmatiene in Viziunea ActualaVasilii90Încă nu există evaluări
- Revista Bistritei X XI 1997Document340 paginiRevista Bistritei X XI 1997CIMECÎncă nu există evaluări
- Cursuri Petrologie Sedimentara Anul IIDocument159 paginiCursuri Petrologie Sedimentara Anul IIIonutÎncă nu există evaluări
- Grecu PDFDocument8 paginiGrecu PDFPop Vasile ViorelÎncă nu există evaluări
- Morfologia Oc. OlanetarDocument12 paginiMorfologia Oc. OlanetarGyu LyaÎncă nu există evaluări
- Codru MomaDocument30 paginiCodru MomaIoan PascuÎncă nu există evaluări
- Elemente de monografie ale comunei Alunis din judetul ClujDe la EverandElemente de monografie ale comunei Alunis din judetul ClujEvaluare: 5 din 5 stele5/5 (1)