S-ar putea să vă placă și
- Test Botanică Examen Biol. FR 2016 - RezDocument10 paginiTest Botanică Examen Biol. FR 2016 - RezMihaela Ivan100% (1)
- ApasiiDocument34 paginiApasiiAlex BejenaruÎncă nu există evaluări
- CURS 11 Formarea Gametofit. FEM SI MASCDocument28 paginiCURS 11 Formarea Gametofit. FEM SI MASCSimona PîslaruÎncă nu există evaluări
- Etologie - Comportament LupDocument16 paginiEtologie - Comportament Lupgabriela serban100% (1)
- Temă Fitopatologie SimptomeDocument2 paginiTemă Fitopatologie SimptomeAndra-Amalia ArmulescuÎncă nu există evaluări
- Zoologia NevertebratelorDocument209 paginiZoologia NevertebratelorRaluca GuriencuÎncă nu există evaluări
- LP1-protectia MunciiDocument1 paginăLP1-protectia MunciiAndra-Amalia ArmulescuÎncă nu există evaluări
- Zoologia NevertebratelorDocument209 paginiZoologia NevertebratelorRaluca GuriencuÎncă nu există evaluări
- Particularităţi Ale Vederii La InsecteDocument7 paginiParticularităţi Ale Vederii La Insectedragos1992Încă nu există evaluări
- Fisa de Lucru LacDocument3 paginiFisa de Lucru LacDaniela AvramÎncă nu există evaluări
- Meioza - Diviziunea MeioticaDocument13 paginiMeioza - Diviziunea MeioticaIoana AgafieiÎncă nu există evaluări
- Referat Arahnide - Cls 6Document2 paginiReferat Arahnide - Cls 6Mirela MarinescuÎncă nu există evaluări
- MuschiDocument3 paginiMuschiMihaela CojocaruÎncă nu există evaluări
- Curs 5Document55 paginiCurs 5Daniela DanitzuÎncă nu există evaluări
- Algele, Oomicetele Și Mixomicetele.Document16 paginiAlgele, Oomicetele Și Mixomicetele.Maria 25kÎncă nu există evaluări
- Ciuperci ToxiceDocument16 paginiCiuperci ToxiceEugen PiveniÎncă nu există evaluări
- PasarileDocument3 paginiPasarileDan GabiÎncă nu există evaluări
- Încrengătura Pteridophyta: A. Ferigile Sunt Primele Plante Vasculare Apărute PeDocument3 paginiÎncrengătura Pteridophyta: A. Ferigile Sunt Primele Plante Vasculare Apărute Peniculinavleju9853Încă nu există evaluări
- ReptileDocument19 paginiReptilebica_alin_adrianÎncă nu există evaluări
- Regnul Animal Proiect BiologieDocument60 paginiRegnul Animal Proiect BiologieAlex BuruianaÎncă nu există evaluări
- LicentaDocument17 paginiLicentaDiana MariaÎncă nu există evaluări
- Genetica Umana Clasa A IX-aDocument5 paginiGenetica Umana Clasa A IX-atudoracheconstantaÎncă nu există evaluări
- Plantele Superioare Cu Spori 10Document11 paginiPlantele Superioare Cu Spori 10Aurelia TarnaÎncă nu există evaluări
- Pasari Picioroange Si AgatatoareDocument2 paginiPasari Picioroange Si AgatatoareEmilutza VladutÎncă nu există evaluări
- Fisa de Activitate BacteriiDocument2 paginiFisa de Activitate BacteriiValentina HîncuÎncă nu există evaluări
- Retile PPVDocument40 paginiRetile PPVNadea BircaÎncă nu există evaluări
- Domesticirea Animalelor DomesticeDocument2 paginiDomesticirea Animalelor Domesticeaxinte2015Încă nu există evaluări
- OrnitorinculDocument11 paginiOrnitorinculalexandruniga88Încă nu există evaluări
- PasariDocument12 paginiPasariLozanu VeraÎncă nu există evaluări
- Hibridarea Si ConsangvinizareaDocument9 paginiHibridarea Si ConsangvinizareaAndreea ElenaÎncă nu există evaluări
- Curs 6Document14 paginiCurs 6Mihaela AlexandraÎncă nu există evaluări
- Zoologia NevertebratelorDocument25 paginiZoologia NevertebratelorLaurentiu BaiuÎncă nu există evaluări
- Regnul AnimaliaDocument25 paginiRegnul AnimaliaMirela ArdeleanÎncă nu există evaluări
- Genetică Mendeliană IDocument53 paginiGenetică Mendeliană IAnda OajdeaÎncă nu există evaluări
- Floarea AlcatuireDocument18 paginiFloarea AlcatuireHamlet Petronius100% (1)
- Fructe Si SeminteDocument8 paginiFructe Si SeminteGabriel Tomac0% (1)
- Ecosisteme Din Alte Zone Ale GlobuluiDocument2 paginiEcosisteme Din Alte Zone Ale GlobuluiValentina BudurcaÎncă nu există evaluări
- Articulatiile MembrelorDocument3 paginiArticulatiile MembrelorTepeş-Stroia EugenÎncă nu există evaluări
- Curs NevertebrateDocument123 paginiCurs NevertebrateEugen VoloșciucÎncă nu există evaluări
- Alimentatia PasarilorDocument12 paginiAlimentatia PasarilorIulian GhismanÎncă nu există evaluări
- Indici EcologiciDocument13 paginiIndici Ecologicivladutualina100% (1)
- Varanul de KomodoDocument13 paginiVaranul de KomodoAdelina PopaÎncă nu există evaluări
- Antilopa ElanDocument1 paginăAntilopa ElanCarla BuiciagÎncă nu există evaluări
- 1 LicheniiDocument11 pagini1 LicheniiVICTORIAÎncă nu există evaluări
- FerigaDocument3 paginiFerigaAdelina LorenaÎncă nu există evaluări
- Reticul EndoplasmaticDocument3 paginiReticul EndoplasmaticAnda AchimÎncă nu există evaluări
- Reptile LeDocument17 paginiReptile Ledr4605Încă nu există evaluări
- Muschiul de PamantDocument9 paginiMuschiul de PamantRodica RussuÎncă nu există evaluări
- Manual Clasa A IX-A Domeniile BiologieiDocument44 paginiManual Clasa A IX-A Domeniile BiologieiMIHAIÎncă nu există evaluări
- 1 ProtozoareDocument19 pagini1 ProtozoarecdyanaÎncă nu există evaluări
- Miscarile PlantelorDocument23 paginiMiscarile PlantelorTatiana NederitaÎncă nu există evaluări
- Câine PDFDocument4 paginiCâine PDFANGHELÎncă nu există evaluări
- Tesuturile 2013Document76 paginiTesuturile 2013Marius PleseaÎncă nu există evaluări
- Entomologie SemDocument30 paginiEntomologie SemIoana Raluca CÎncă nu există evaluări
- Celula EucariotaDocument16 paginiCelula EucariotaMirela ArdeleanÎncă nu există evaluări
- PăsăriDocument52 paginiPăsăriAlexandru Mihai Mihai Alexandru Cucu100% (1)
- Locomoția La Seria AnimalăDocument10 paginiLocomoția La Seria AnimalăRoxanaÎncă nu există evaluări
- Biologie Recap FilanaDocument11 paginiBiologie Recap FilanaTărsănaru TudorÎncă nu există evaluări
- c10. Transmiterea Caracterelor La EucarioteDocument38 paginic10. Transmiterea Caracterelor La EucarioteGeorgiana Daniela Dragomir100% (1)
- Genetica MicroorganismelorDocument2 paginiGenetica MicroorganismelorAna-Maria LopotencoÎncă nu există evaluări
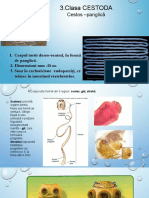
- Clasa CestodaDocument51 paginiClasa CestodaEmmaÎncă nu există evaluări
- Lucrare Diploma Evolutie Si Tratament in AvcDocument26 paginiLucrare Diploma Evolutie Si Tratament in AvcGabriela GabÎncă nu există evaluări
- SISTEMUL Nervos - Docc8a30Document7 paginiSISTEMUL Nervos - Docc8a30Lucia RosuÎncă nu există evaluări
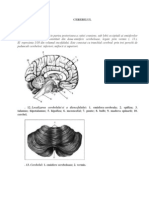
- CEREBELULDocument14 paginiCEREBELULsimiusaghiÎncă nu există evaluări
- Tema 1 LP FiziopatologieDocument1 paginăTema 1 LP FiziopatologieAndra-Amalia ArmulescuÎncă nu există evaluări
- Ecomorfologia Plantelor ResearchGateDocument20 paginiEcomorfologia Plantelor ResearchGateAndra-Amalia ArmulescuÎncă nu există evaluări
- Cursuri FV 1 - 191Document130 paginiCursuri FV 1 - 191Andra-Amalia ArmulescuÎncă nu există evaluări
- R.Ag-Ac - Etape Si ParticDocument4 paginiR.Ag-Ac - Etape Si ParticAndra-Amalia ArmulescuÎncă nu există evaluări
- Curs SNCDocument36 paginiCurs SNCAndra-Amalia ArmulescuÎncă nu există evaluări
- Curs Transm Sin 2016Document45 paginiCurs Transm Sin 2016Andra-Amalia ArmulescuÎncă nu există evaluări
- Subiecte Examen Biologie Celulara 2019Document3 paginiSubiecte Examen Biologie Celulara 2019Andra-Amalia ArmulescuÎncă nu există evaluări
- Curs 1Document2 paginiCurs 1Andra-Amalia ArmulescuÎncă nu există evaluări
- Formular de Inscriere Ca Voluntar 2013Document3 paginiFormular de Inscriere Ca Voluntar 2013Octavian PopoviciÎncă nu există evaluări
- Subiecte Grila 2019Document14 paginiSubiecte Grila 2019Andra-Amalia Armulescu100% (1)
- Subiecte Examen Biologie Celulara 2019Document3 paginiSubiecte Examen Biologie Celulara 2019Andra-Amalia ArmulescuÎncă nu există evaluări
- PILDADocument1 paginăPILDAAndra-Amalia ArmulescuÎncă nu există evaluări
- Fiziologie Vegetala c1 c2Document11 paginiFiziologie Vegetala c1 c2Mihaila OanaÎncă nu există evaluări
- MiologieDocument13 paginiMiologieAndra-Amalia ArmulescuÎncă nu există evaluări
- Teste Biologie RODocument17 paginiTeste Biologie ROMateescu Cristina100% (4)
- Pilda 22Document1 paginăPilda 22Andra-Amalia ArmulescuÎncă nu există evaluări
- Pilda 22Document1 paginăPilda 22Andra-Amalia ArmulescuÎncă nu există evaluări
- Probleme Rezolvate de Statistica Matematica - 2017 - 2018Document27 paginiProbleme Rezolvate de Statistica Matematica - 2017 - 2018Andra-Amalia ArmulescuÎncă nu există evaluări
- Pilda 22Document1 paginăPilda 22Andra-Amalia ArmulescuÎncă nu există evaluări
- Teme Referate La Disciplina ADSEDocument2 paginiTeme Referate La Disciplina ADSEAndra-Amalia ArmulescuÎncă nu există evaluări
- Seminarul 4 Mate in Bio 2017 - 2018Document9 paginiSeminarul 4 Mate in Bio 2017 - 2018Andra-Amalia ArmulescuÎncă nu există evaluări
- Seminarul 2 Mat in Bio 2017 - 2018Document12 paginiSeminarul 2 Mat in Bio 2017 - 2018Andra-Amalia ArmulescuÎncă nu există evaluări
- Evaluarea Proiectelor de InvestitiiDocument13 paginiEvaluarea Proiectelor de Investitiistf2xÎncă nu există evaluări
- Responsabili de CatedraDocument1 paginăResponsabili de CatedraAndra-Amalia ArmulescuÎncă nu există evaluări
- Performanta IntreprinderiiDocument1 paginăPerformanta IntreprinderiiAndra-Amalia ArmulescuÎncă nu există evaluări