S-ar putea să vă placă și
- Sistemul Circulator La VertebrateDocument1 paginăSistemul Circulator La VertebrateVic AlÎncă nu există evaluări
- Structura Si Functiile Sistemului Nervos CentralDocument7 paginiStructura Si Functiile Sistemului Nervos CentralAlexGRÎncă nu există evaluări
- Forme de Manifestare A ForteiDocument3 paginiForme de Manifestare A Forteimarius dimaÎncă nu există evaluări
- Curs 1 - ReceptoriDocument5 paginiCurs 1 - ReceptoriAura ConstantinÎncă nu există evaluări
- Clasa AmphibiaDocument33 paginiClasa AmphibiaMihneaÎncă nu există evaluări
- Se Acorda 1 Punct Din Oficiu!Document1 paginăSe Acorda 1 Punct Din Oficiu!Ileana Grigore100% (1)
- Absorbtia IntestinalaDocument2 paginiAbsorbtia IntestinalaFlorentina TerghesÎncă nu există evaluări
- Plămânii - Topografie, Structură Şi Funcţii. Mecanismul Respiraţiei.Document15 paginiPlămânii - Topografie, Structură Şi Funcţii. Mecanismul Respiraţiei.Carolina HîncuÎncă nu există evaluări
- Structura ProteinelorDocument13 paginiStructura ProteinelorA B100% (1)
- Parcul National Muntii RodneiDocument4 paginiParcul National Muntii RodneiApostol AlexandruÎncă nu există evaluări
- Ghid Admitere Licenta Efs 2019-SITEDocument54 paginiGhid Admitere Licenta Efs 2019-SITEAlina GeorgianaÎncă nu există evaluări
- Citoplasma CelularaDocument16 paginiCitoplasma CelularaEnea DanielaÎncă nu există evaluări
- Muschii CoxaliDocument2 paginiMuschii CoxaliAliceÎncă nu există evaluări
- Test Aparatul LocomotorDocument2 paginiTest Aparatul LocomotorAndreea GhițăÎncă nu există evaluări
- Mecanismul DigestieiDocument17 paginiMecanismul Digestieidoruk1964Încă nu există evaluări
- Orar Liceul GH Ruset Roznovanu Dec 2017Document50 paginiOrar Liceul GH Ruset Roznovanu Dec 2017Grup Scolar RoznovÎncă nu există evaluări
- Functiile Fundamentale Ale OrganismuluiDocument3 paginiFunctiile Fundamentale Ale OrganismuluiBeatrice ChiurciuÎncă nu există evaluări
- Sistemul Digestiv La MamifereDocument21 paginiSistemul Digestiv La MamifereIoana Trif100% (1)
- Aparatul VestibularDocument17 paginiAparatul VestibularStud2345Încă nu există evaluări
- Biochimie Curs 1 Anu 1 Sem 2Document6 paginiBiochimie Curs 1 Anu 1 Sem 2Roxana Nae PuccaÎncă nu există evaluări
- Organizarea Structurala Si Functionala A CeluleiDocument19 paginiOrganizarea Structurala Si Functionala A CeluleiGiuliaGoth00Încă nu există evaluări
- Comparatie Intre SNV Simpatic Si SNV ParasimpaticDocument2 paginiComparatie Intre SNV Simpatic Si SNV ParasimpaticSteffNedelcuÎncă nu există evaluări
- ACTIVITATEA NERVOASĂ SUPERIOAraDocument8 paginiACTIVITATEA NERVOASĂ SUPERIOAraAda StilleÎncă nu există evaluări
- Caile de Conducere Ale SNCDocument18 paginiCaile de Conducere Ale SNCAndreea GheorghiuÎncă nu există evaluări
- Funcția Autocatalitică Și HeterocataliticăDocument6 paginiFuncția Autocatalitică Și HeterocataliticăM2thylÎncă nu există evaluări
- Tesutul Muscular Scheletic - BFK - 2022 PDFDocument45 paginiTesutul Muscular Scheletic - BFK - 2022 PDFLorena StroeÎncă nu există evaluări
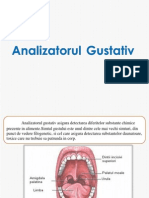
- Analizatorul GustativDocument11 paginiAnalizatorul GustativCristina NasaleanÎncă nu există evaluări
- Celula FiziologieDocument33 paginiCelula FiziologieClaudiu Boarţă100% (1)
- Epitelii de Acoepitelii de Acoperire - PptperireDocument40 paginiEpitelii de Acoepitelii de Acoperire - PptperireAnonymous 51EI0pÎncă nu există evaluări
- Prezentare AnticorpiDocument15 paginiPrezentare AnticorpiOanaDorofteÎncă nu există evaluări
- Calitatile Motrice Sunt Insusiri Ale Organismului Care Ofera Individului Posibilitatea Executarii Diferitelor Acte MotriceDocument9 paginiCalitatile Motrice Sunt Insusiri Ale Organismului Care Ofera Individului Posibilitatea Executarii Diferitelor Acte MotriceIonut MitreaÎncă nu există evaluări
- Tipuri de Contractii MusculareDocument4 paginiTipuri de Contractii MusculareNelu Platon100% (1)
- Fiziologia Analizatorului Vestibular Utricula Sacula Şi Canalele SemicirculareDocument1 paginăFiziologia Analizatorului Vestibular Utricula Sacula Şi Canalele SemicirculareDiana LavricÎncă nu există evaluări
- SISTEMUL LOCOMOTOR LA MAMIFERE, Sinteză, Cl. A X ADocument7 paginiSISTEMUL LOCOMOTOR LA MAMIFERE, Sinteză, Cl. A X AHăpăianu ElisaÎncă nu există evaluări
- Intrebari AnatomieDocument124 paginiIntrebari AnatomieVasile LozinschiiÎncă nu există evaluări
- Mecanismul Formării UrineiDocument2 paginiMecanismul Formării UrineiPintilie RalucaÎncă nu există evaluări
- StructuraDocument9 paginiStructuraInga FulgerÎncă nu există evaluări
- Echilibre Si Dezechilibre NaturaleDocument10 paginiEchilibre Si Dezechilibre NaturaleIulia Vintilă100% (1)
- Absorbţia Apei de Către PlanteDocument2 paginiAbsorbţia Apei de Către PlanteV CÎncă nu există evaluări
- Fiziologia Muschilor StriatiDocument8 paginiFiziologia Muschilor Striatinigor1297Încă nu există evaluări
- 45899-19046 Viermi CilindriciDocument9 pagini45899-19046 Viermi CilindriciDariusGeniusÎncă nu există evaluări
- Legatura IonicaDocument7 paginiLegatura IonicaDomnica SapojnicÎncă nu există evaluări
- 12.REFERAT-Fauna Apelor CurgatoareDocument7 pagini12.REFERAT-Fauna Apelor CurgatoareBistea Iuliana ElenaÎncă nu există evaluări
- Regulament Olimpiada de Pregatire Sportiva TeoreticaDocument13 paginiRegulament Olimpiada de Pregatire Sportiva TeoreticaAvram NeluÎncă nu există evaluări
- SurfingulDocument6 paginiSurfingulMihaela Geanina BădinÎncă nu există evaluări
- Rolul Campaniilor de Promovare A Unui Stil de ViatDocument7 paginiRolul Campaniilor de Promovare A Unui Stil de Viatmarieanne984everÎncă nu există evaluări
- Totul Despre ProteineDocument9 paginiTotul Despre ProteineCostea MariaÎncă nu există evaluări
- Analizatorul VizualDocument6 paginiAnalizatorul VizualIrina ASÎncă nu există evaluări
- Structura StomaculuiDocument3 paginiStructura StomaculuiCebotari IvanÎncă nu există evaluări
- Planificare Biologie Clasa A VIII-A - ModelDocument4 paginiPlanificare Biologie Clasa A VIII-A - ModelSabina CurcaÎncă nu există evaluări
- Variante Romana Partea I Rezolvate 2009 M EminescuDocument10 paginiVariante Romana Partea I Rezolvate 2009 M EminescuElyzu0% (1)
- Fiziologia Fibrei MusculareDocument52 paginiFiziologia Fibrei MusculareNesrin Bactali100% (5)
- Curs Fiziologia Mușchilor Striați Și Netezi-34826Document57 paginiCurs Fiziologia Mușchilor Striați Și Netezi-34826Alina IlovanÎncă nu există evaluări
- Fiziologia Fibrei MusculareDocument52 paginiFiziologia Fibrei MusculareDaniel HuntelarÎncă nu există evaluări
- 1.5. Fiziologia Fibrei MusculareDocument82 pagini1.5. Fiziologia Fibrei MusculareAlina Topor Psi-MedicalÎncă nu există evaluări
- Curs - Muschi StriatDocument40 paginiCurs - Muschi StriatDiana Didi Dia100% (1)
- Cursul 4 FizioDocument5 paginiCursul 4 Fiziodanyela0808Încă nu există evaluări
- MuscularDocument9 paginiMuscularpufdepapadieÎncă nu există evaluări
- Sistemul NeuromuscDocument67 paginiSistemul NeuromuscAlexandru PurcelÎncă nu există evaluări
- Curs 4 Login 2014Document5 paginiCurs 4 Login 2014Ema MocanÎncă nu există evaluări
- Florile DalbeDocument1 paginăFlorile DalbeOana CristinaÎncă nu există evaluări
- UprinsDocument28 paginiUprinsOana CristinaÎncă nu există evaluări
- Povestea Lui Moş CrăciunDocument3 paginiPovestea Lui Moş CrăciunOana CristinaÎncă nu există evaluări
- Astazi S-A Nascut HristosDocument1 paginăAstazi S-A Nascut HristosOana CristinaÎncă nu există evaluări
- Cerul Si PamantulDocument2 paginiCerul Si PamantulOana CristinaÎncă nu există evaluări
- Colin DitaDocument1 paginăColin DitaOana CristinaÎncă nu există evaluări
- Duminica 29 Dupa RusaliiDocument1 paginăDuminica 29 Dupa RusaliiOana CristinaÎncă nu există evaluări
- 2 - CURS - CTG Si Supravegherea Perfuziei Cu OxitonDocument17 pagini2 - CURS - CTG Si Supravegherea Perfuziei Cu OxitonOana CristinaÎncă nu există evaluări
- 4 - CURS - Operatia CezarianaDocument28 pagini4 - CURS - Operatia CezarianaOana CristinaÎncă nu există evaluări
- 3 - CURS - Lehuzia FiziologicaDocument22 pagini3 - CURS - Lehuzia FiziologicaOana CristinaÎncă nu există evaluări
- Patologia Benignă A Sânului, Cancer de SânDocument32 paginiPatologia Benignă A Sânului, Cancer de SânOana Cristina100% (2)
- 1 - CURS - Supravegherea Parturientei. Monitorizarea CU. BCFDocument45 pagini1 - CURS - Supravegherea Parturientei. Monitorizarea CU. BCFOana Cristina100% (2)
- EndometriozaDocument9 paginiEndometriozaOana Cristina100% (2)