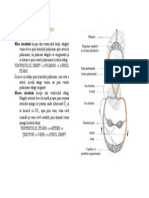
S-ar putea să vă placă și
- Odiseea genomului: Mistere medicale și aventura incredibilă a dezlegării lorDe la EverandOdiseea genomului: Mistere medicale și aventura incredibilă a dezlegării lorÎncă nu există evaluări
- Microcirculația sanguină: Următoarea generație de asistență socialăDe la EverandMicrocirculația sanguină: Următoarea generație de asistență socialăÎncă nu există evaluări
- Referat CromozomiDocument12 paginiReferat CromozomiCorina Ioana100% (1)
- Suport Didactic La Biologie Clasa IXDocument39 paginiSuport Didactic La Biologie Clasa IXMihai Boinceanu2004100% (2)
- Referat CromozomiDocument11 paginiReferat CromozomiIonuț Cristian Mihoci80% (5)
- Mitoza Si MeiozaDocument8 paginiMitoza Si MeiozaAnastasia Frinea100% (4)
- Curs III Management 2017Document35 paginiCurs III Management 2017Ungureanu MikeÎncă nu există evaluări
- Diviziunea CelularaDocument21 paginiDiviziunea CelularaSareh Mehran100% (2)
- Asemanari Si Deoseb Celule Proc. EucDocument2 paginiAsemanari Si Deoseb Celule Proc. EucAdelle Bio100% (1)
- Structura Cromozomilor Si Categorii de CromozomiDocument9 paginiStructura Cromozomilor Si Categorii de CromozomiConstantinescu AnamariaÎncă nu există evaluări
- Diviziunea CelularăDocument7 paginiDiviziunea Celularăcrisss_tinaÎncă nu există evaluări
- Diviziunea CelularaDocument4 paginiDiviziunea CelularaAle XaÎncă nu există evaluări
- Curs Genetica MVDocument55 paginiCurs Genetica MVaa_alexw_aaÎncă nu există evaluări
- Structura CromozomuluiDocument12 paginiStructura CromozomuluiAnca Mandache100% (1)
- Cum Se Nasc CeluleleDocument3 paginiCum Se Nasc CeluleleMathew Arnold67% (3)
- CromozomiiDocument4 paginiCromozomiiDorina LucaÎncă nu există evaluări
- Analiza Swot FermăDocument5 paginiAnaliza Swot FermăUngureanu MikeÎncă nu există evaluări
- Bazele Citologice Ale EreditățiiDocument25 paginiBazele Citologice Ale EreditățiiAna Suruceanu0% (1)
- Diviziunea Celulara Mitoza Si MeiozaDocument10 paginiDiviziunea Celulara Mitoza Si Meiozazaga100% (1)
- Diviziunea CelularăDocument7 paginiDiviziunea CelularăGeorge BrinzeaÎncă nu există evaluări
- Curs GeneticaDocument20 paginiCurs GeneticanicoletaÎncă nu există evaluări
- Plan de Afaceri Salon de InfrumusetareDocument13 paginiPlan de Afaceri Salon de InfrumusetareUngureanu MikeÎncă nu există evaluări
- MeiozaDocument4 paginiMeiozaLaurentiu BaiuÎncă nu există evaluări
- Bruceloza 2Document14 paginiBruceloza 2Ungureanu MikeÎncă nu există evaluări
- Mitoza PDFDocument78 paginiMitoza PDFRazvan Luca100% (2)
- Cromozomii 22Document18 paginiCromozomii 22Ama PredaÎncă nu există evaluări
- CromozomDocument5 paginiCromozomDana CozmaÎncă nu există evaluări
- Biocel LP 9-Mitoza PDFDocument30 paginiBiocel LP 9-Mitoza PDFOana RaduÎncă nu există evaluări
- Interfaza Si Diviziunea CelularaDocument11 paginiInterfaza Si Diviziunea CelularaAdriana Crăciun50% (2)
- LicentaDocument4 paginiLicentaUngureanu MikeÎncă nu există evaluări
- Mica Si Marea CirculatieDocument1 paginăMica Si Marea CirculatieUngureanu MikeÎncă nu există evaluări
- Genetic ADocument16 paginiGenetic AUngureanu MikeÎncă nu există evaluări
- Tema 5 CromosomiiDocument9 paginiTema 5 CromosomiiOxana SoltanÎncă nu există evaluări
- CromozomiiDocument3 paginiCromozomiiSabina SerbuÎncă nu există evaluări
- Structura CromozomilorDocument23 paginiStructura CromozomilorCorina IoanaÎncă nu există evaluări
- Genetica Citologica ÅDocument4 paginiGenetica Citologica Åolguta-mihaela.petre.fpÎncă nu există evaluări
- Diviziunea Celulara - Curs 4Document9 paginiDiviziunea Celulara - Curs 4Adrian MorosanÎncă nu există evaluări
- BiologiaDocument36 paginiBiologiaDubalariNicolaiÎncă nu există evaluări
- Diviziunea CelularaDocument21 paginiDiviziunea CelularaTiberiu MindrescuÎncă nu există evaluări
- LP6 Diviziunea CelularaDocument8 paginiLP6 Diviziunea CelularaMaria MuresanÎncă nu există evaluări
- GENOMICA CURS 7 Structura Internă A Cromosomului EucariotDocument11 paginiGENOMICA CURS 7 Structura Internă A Cromosomului EucariotCristina ButucÎncă nu există evaluări
- Embriologie ReferatDocument13 paginiEmbriologie ReferatElena Alexandra Beladan100% (1)
- Referat docxVIOLETADocument10 paginiReferat docxVIOLETAtuturici liviuÎncă nu există evaluări
- Diviziunea CelularaDocument9 paginiDiviziunea CelularaAlexander CrowlordÎncă nu există evaluări
- Embriologie Curs 11-Genom - Cromoz.cod - Gen Si Det - Gr.sg.Document11 paginiEmbriologie Curs 11-Genom - Cromoz.cod - Gen Si Det - Gr.sg.Alexandru GiubegaÎncă nu există evaluări
- Biologie Clasa A 9aDocument3 paginiBiologie Clasa A 9aPlopik EdyÎncă nu există evaluări
- 03 Alcatuirea Cariotipului La PlanteDocument5 pagini03 Alcatuirea Cariotipului La PlanteAlex CarpÎncă nu există evaluări
- Diviziunea Celulara A Celulei Normale EucarioteDocument3 paginiDiviziunea Celulara A Celulei Normale EucarioteSirb AlexandruÎncă nu există evaluări
- WWW - Referat.ro Meioza - Docf80fdDocument6 paginiWWW - Referat.ro Meioza - Docf80fdsweet_roxy29927569Încă nu există evaluări
- Diviziunea CelularaDocument26 paginiDiviziunea CelularaAnca DragoiÎncă nu există evaluări
- Curs 2 Biol. MolecularaDocument5 paginiCurs 2 Biol. MolecularaLaura NeaguÎncă nu există evaluări
- MitozaDocument19 paginiMitozaIoana RoxanaÎncă nu există evaluări
- Genotipul Uman PatologicDocument8 paginiGenotipul Uman PatologicRuxi CiaglicÎncă nu există evaluări
- Biologie MolecularaDocument29 paginiBiologie MolecularaMariana GhermanÎncă nu există evaluări
- GametogenezaDocument29 paginiGametogenezaMaria RotarÎncă nu există evaluări
- Diviz CelularaDocument26 paginiDiviz CelularaAbrudan PatriciaÎncă nu există evaluări
- Diviziunea CelularaDocument9 paginiDiviziunea CelularaDurau ElizaÎncă nu există evaluări
- DiviziuneaDocument29 paginiDiviziuneaVascovici DanÎncă nu există evaluări
- BioDocument4 paginiBioSarahÎncă nu există evaluări
- Cap1 (Mitoza)Document41 paginiCap1 (Mitoza)crindefloareÎncă nu există evaluări
- Referat BiologieDocument6 paginiReferat BiologieCucu OleseaÎncă nu există evaluări
- Снимок экрана 2023-10-01 в 15.50.57Document39 paginiСнимок экрана 2023-10-01 в 15.50.57Максим БулинаÎncă nu există evaluări
- Curs Ii Management 2017Document21 paginiCurs Ii Management 2017Ungureanu MikeÎncă nu există evaluări
- Curs I Management 2017Document18 paginiCurs I Management 2017Ungureanu MikeÎncă nu există evaluări
- Curs 7 SP DigDocument105 paginiCurs 7 SP DigUngureanu MikeÎncă nu există evaluări
- Asandei Adina MV 2018Document18 paginiAsandei Adina MV 2018Ungureanu MikeÎncă nu există evaluări
- Curs 6 Gen TumoriDocument23 paginiCurs 6 Gen TumoriUngureanu MikeÎncă nu există evaluări
- Curs 2 Gen MecPatGen InflamatiiDocument98 paginiCurs 2 Gen MecPatGen InflamatiiUngureanu MikeÎncă nu există evaluări
- A6 4 Anexa 2 Sablon Plan Afaceri ConcursDocument20 paginiA6 4 Anexa 2 Sablon Plan Afaceri ConcursUngureanu MikeÎncă nu există evaluări
- Titluri Desene AnatomieDocument2 paginiTitluri Desene AnatomieUngureanu MikeÎncă nu există evaluări
- BiofizicaDocument7 paginiBiofizicaUngureanu MikeÎncă nu există evaluări
- Simptomele Intoxicaţiei Cu TTX Încep La Câteva Minute După Ingerarea SubstanţeiDocument8 paginiSimptomele Intoxicaţiei Cu TTX Încep La Câteva Minute După Ingerarea SubstanţeiUngureanu MikeÎncă nu există evaluări