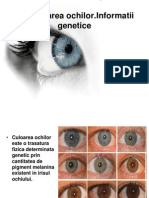
S-ar putea să vă placă și
- Războiul sexelor. Infidelitate, conflicte sexuale și alte lupte din dormitorDe la EverandRăzboiul sexelor. Infidelitate, conflicte sexuale și alte lupte din dormitorEvaluare: 3.5 din 5 stele3.5/5 (2)
- Mutatiile Cromozomiale Importanta PracticaDocument18 paginiMutatiile Cromozomiale Importanta PracticaStefan Alexandru100% (2)
- Structura Analiza Şi Localizarea GenelorDocument48 paginiStructura Analiza Şi Localizarea GenelorautorulaneiÎncă nu există evaluări
- Cap 7 Ameliorarea Genetica A Familiilor de AlbineDocument31 paginiCap 7 Ameliorarea Genetica A Familiilor de AlbineMaria-Madalina Bodescu100% (1)
- Eco Genomic ADocument8 paginiEco Genomic AradulusÎncă nu există evaluări
- Alele GeneDocument8 paginiAlele GeneIulia Vasilica GirosanÎncă nu există evaluări
- Mostenirea Caracterelor GenelorDocument11 paginiMostenirea Caracterelor GenelorAndrian CusmirÎncă nu există evaluări
- GeneleDocument6 paginiGeneleConstantin Gonta100% (1)
- Referat Transl. Rob.-Kiss ÁgnesDocument8 paginiReferat Transl. Rob.-Kiss ÁgnesAgnes Teodora KissÎncă nu există evaluări
- Teoria Cromozomiala A EreditatiiDocument55 paginiTeoria Cromozomiala A EreditatiiKent EdwardsÎncă nu există evaluări
- Genetica - Genetica PopulationalaDocument5 paginiGenetica - Genetica PopulationalaAnca TeodoroiuÎncă nu există evaluări
- S8 Embriologie Si Genetica CursDocument2 paginiS8 Embriologie Si Genetica Cursrosse_maryÎncă nu există evaluări
- Articol Despre GemeniDocument3 paginiArticol Despre GemeniSorana MoleaÎncă nu există evaluări
- Psihologia VârstelorDocument118 paginiPsihologia VârstelorEhh EnNaÎncă nu există evaluări
- Unitatea 1 Legile MendelieneDocument26 paginiUnitatea 1 Legile MendelieneAna Maria MacrescuÎncă nu există evaluări
- 5 Antropologie GeneticăDocument14 pagini5 Antropologie GeneticăAnca IoanaÎncă nu există evaluări
- Determinismul Genetic Al SexelorDocument9 paginiDeterminismul Genetic Al SexelorVtZ50% (4)
- PRENATAL SI Nou NascutulDocument16 paginiPRENATAL SI Nou NascutulElena Dediu100% (2)
- Sarcina MultiplaDocument4 paginiSarcina MultiplaAnk GeorgyÎncă nu există evaluări
- Tema 9.genetica SexuluiDocument11 paginiTema 9.genetica SexuluisilviaÎncă nu există evaluări
- Cap 3 Gene Și Gene de ComportamentDocument32 paginiCap 3 Gene Și Gene de ComportamentValentin SinÎncă nu există evaluări
- Clonarea UmanaDocument8 paginiClonarea UmanaIrina CornilovÎncă nu există evaluări
- 4 Linkage Si AsociereDocument61 pagini4 Linkage Si AsociereMadalina PişteaÎncă nu există evaluări
- WWW - Referat.ro Genetica A81c0Document12 paginiWWW - Referat.ro Genetica A81c0AntociAlexeiÎncă nu există evaluări
- Moştenirea Înlănţuită A CaracterelorDocument7 paginiMoştenirea Înlănţuită A CaracterelorVlaicu TudorÎncă nu există evaluări
- C2 Cromozomi GenaDocument10 paginiC2 Cromozomi GenaAndreea CsoltiÎncă nu există evaluări
- Teoria Cromozomiala A EreditatiiDocument55 paginiTeoria Cromozomiala A EreditatiiSvsu SuseniÎncă nu există evaluări
- ConsangvinizareaDocument14 paginiConsangvinizareaValentina Vorona100% (1)
- Animale TransgeniceDocument13 paginiAnimale TransgeniceElenaDanielaJaleru100% (1)
- Unitatea 6 - Teoria Cromozomiala A EreditatiiDocument11 paginiUnitatea 6 - Teoria Cromozomiala A EreditatiiBucur AlexandruÎncă nu există evaluări
- Teoria Cromozomială A EreditățiiDocument19 paginiTeoria Cromozomială A EreditățiiAlexia PăpădieÎncă nu există evaluări
- Probleme GeneticaDocument9 paginiProbleme GeneticaCosmin Pavel100% (1)
- Recombinarea GeneticăDocument12 paginiRecombinarea Geneticămaria morariuÎncă nu există evaluări
- Testul de Genetică A Populației IIDocument12 paginiTestul de Genetică A Populației IIScribdTranslationsÎncă nu există evaluări
- Diagnostic PrenatalDocument26 paginiDiagnostic PrenatalEmanuela GhilasiÎncă nu există evaluări
- Arbori Genealogici9Document28 paginiArbori Genealogici9DemnitateÎncă nu există evaluări
- Legea Echilibrelor În PopulaţieDocument6 paginiLegea Echilibrelor În PopulaţieIrina FilipÎncă nu există evaluări
- Genetica SibiuDocument57 paginiGenetica SibiuBebe Ionuț AnițaÎncă nu există evaluări
- Mostenirea CaracterelorDocument11 paginiMostenirea CaracterelorIon GrigorasÎncă nu există evaluări
- II GeneticaDocument17 paginiII GeneticaAlina PaşcaÎncă nu există evaluări
- Curs 5 PDFDocument12 paginiCurs 5 PDFElena AngheloiuÎncă nu există evaluări
- Genetica Comportamentului UmanDocument76 paginiGenetica Comportamentului UmanGeorgiana RîmniceanuÎncă nu există evaluări
- Proiect Genetica 121212.Document7 paginiProiect Genetica 121212.Vasilescu CristianÎncă nu există evaluări
- Organisme Modificate GeneticDocument32 paginiOrganisme Modificate GeneticLaura Stoica50% (2)
- Bazele Genetice Ale Comportamentului Sinteze de CursDocument77 paginiBazele Genetice Ale Comportamentului Sinteze de Cursliviaserban100% (2)
- 9 Metode de Studiere A EreditatiiDocument7 pagini9 Metode de Studiere A EreditatiiZatoloșnîi Abigaela-ZiniaÎncă nu există evaluări
- 6 Mostenirea Grupeolor SangvineDocument8 pagini6 Mostenirea Grupeolor SangvineZatoloșnîi Abigaela-ZiniaÎncă nu există evaluări
- Material Lp-Genetica Zoo IIDocument48 paginiMaterial Lp-Genetica Zoo IIFlorin MariuteiÎncă nu există evaluări
- Genetica OchilorDocument27 paginiGenetica OchilorBadea Elena-DeliaÎncă nu există evaluări
- Culoarea OchilorDocument27 paginiCuloarea OchilorMonica CiobanuÎncă nu există evaluări
- Genetica Umană MetodeDocument30 paginiGenetica Umană MetodeLacramioara SeriaÎncă nu există evaluări
- 98 Buletin MetodicDocument22 pagini98 Buletin MetodicCarrie VaughnÎncă nu există evaluări
- Boli Genetice Ereditare - Referat Biologie - Clasa A XII-aDocument3 paginiBoli Genetice Ereditare - Referat Biologie - Clasa A XII-aPoseButterfly67% (3)
- Test Biotehnologie 2Document2 paginiTest Biotehnologie 2Veronica ChirtocaÎncă nu există evaluări
- Odiseea genomului: Mistere medicale și aventura incredibilă a dezlegării lorDe la EverandOdiseea genomului: Mistere medicale și aventura incredibilă a dezlegării lorÎncă nu există evaluări