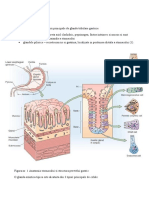
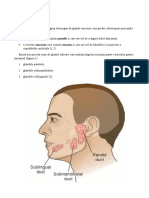
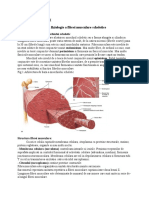
S-ar putea să vă placă și
- Mitoza Si MeiozaDocument8 paginiMitoza Si MeiozaAnastasia Frinea100% (4)
- Curs Genetica Anul IDocument333 paginiCurs Genetica Anul IFlavius Cristuinea100% (2)
- Mitoza Si Meioza - CursDocument18 paginiMitoza Si Meioza - Cursandreeamihai92Încă nu există evaluări
- Curs Genetica MVDocument55 paginiCurs Genetica MVaa_alexw_aaÎncă nu există evaluări
- Diviziune CelularaDocument5 paginiDiviziune CelularaAlexandra LeocaÎncă nu există evaluări
- Ereditatea Monogenica, MultifactDocument10 paginiEreditatea Monogenica, MultifactAga_tthaÎncă nu există evaluări
- Diviziunea CelularaDocument3 paginiDiviziunea CelularaValentinoÎncă nu există evaluări
- Bazele Citologice Ale EreditățiiDocument25 paginiBazele Citologice Ale EreditățiiAna Suruceanu0% (1)
- Interfaza Si Diviziunea CelularaDocument11 paginiInterfaza Si Diviziunea CelularaAdriana Crăciun50% (2)
- MEIOZADocument10 paginiMEIOZAAlexandraS2812Încă nu există evaluări
- 1 - Diviziunea Celulară - CDocument30 pagini1 - Diviziunea Celulară - CCazacu MonicaÎncă nu există evaluări
- Diviziunea CelularăDocument7 paginiDiviziunea CelularăGeorge BrinzeaÎncă nu există evaluări
- Meioza 1Document8 paginiMeioza 1Vlad ConstantinescuÎncă nu există evaluări
- Diviziunea CelularaDocument4 paginiDiviziunea CelularaAle XaÎncă nu există evaluări
- Mitoza PDFDocument78 paginiMitoza PDFRazvan Luca100% (2)
- Gametogeneza Si FecundatiaDocument13 paginiGametogeneza Si FecundatiaCristina-Maria DumitruÎncă nu există evaluări
- 21 Celula - Mitoza - MeiozaDocument5 pagini21 Celula - Mitoza - MeiozaDiana PetreaÎncă nu există evaluări
- Ciclul CelularDocument19 paginiCiclul CelularElena-Catalina Popa100% (1)
- Fiziologie LP NR 6 Anul IDocument17 paginiFiziologie LP NR 6 Anul IAnna MariaÎncă nu există evaluări
- Ciclul CelularDocument25 paginiCiclul CelularAndreea Valcu100% (3)
- 7.5. Proiect Didactic (Sablon)Document6 pagini7.5. Proiect Didactic (Sablon)Valrtina CalinÎncă nu există evaluări
- Meioză Si MitozaDocument5 paginiMeioză Si MitozaDima Botnari100% (1)
- LP6 Diviziunea CelularaDocument8 paginiLP6 Diviziunea CelularaMaria MuresanÎncă nu există evaluări
- GametogenezaDocument29 paginiGametogenezaMaria RotarÎncă nu există evaluări
- LP Nr. 7 Biologie CelularăDocument6 paginiLP Nr. 7 Biologie CelularăDarius FlorentiuÎncă nu există evaluări
- Prezentare 90Document8 paginiPrezentare 90Ana Maria PăunaşÎncă nu există evaluări
- 0Document6 pagini0Zamfirescu Vladimir-AlexandruÎncă nu există evaluări
- MeiozăDocument3 paginiMeiozăMinodora IsarÎncă nu există evaluări
- Studiul Meiozei La OmDocument8 paginiStudiul Meiozei La OmAndra IsabelaÎncă nu există evaluări
- Anatomia Si Fiziologia Omului. Curs 3 - MITOZA, MEIOZADocument5 paginiAnatomia Si Fiziologia Omului. Curs 3 - MITOZA, MEIOZAdinculescu-koborzan constantinÎncă nu există evaluări
- Mitoza Si MeiozaDocument6 paginiMitoza Si MeiozaAleX 2.0Încă nu există evaluări
- LP - 4 - Genetica Medicala - Anul Ii - MDDocument26 paginiLP - 4 - Genetica Medicala - Anul Ii - MDAndra CrisÎncă nu există evaluări
- MeiozaDocument5 paginiMeiozaRadu BasamacÎncă nu există evaluări
- MitozaDocument2 paginiMitozaAndrei-Constantin RosuÎncă nu există evaluări
- 5#pag 85-98 Cu BTDocument14 pagini5#pag 85-98 Cu BTMadalina ChirilaÎncă nu există evaluări
- Comparația Între Mitoză Și MeiozăDocument14 paginiComparația Între Mitoză Și MeiozăAndreea GabrielaÎncă nu există evaluări
- Studiul Meiozei La OmDocument5 paginiStudiul Meiozei La OmFlavius Ion NicolaeÎncă nu există evaluări
- MitozăDocument14 paginiMitozăGiulieta RaileanÎncă nu există evaluări
- WWW - Referat.ro Meioza - Docf80fdDocument6 paginiWWW - Referat.ro Meioza - Docf80fdsweet_roxy29927569Încă nu există evaluări
- Diviziunea CelularăDocument3 paginiDiviziunea CelularăEllenaGuguilaÎncă nu există evaluări
- New Microsoft Word Document 14Document3 paginiNew Microsoft Word Document 14cristiana.gabriela95Încă nu există evaluări
- CURS 7 Biologie CelularaDocument17 paginiCURS 7 Biologie CelularaElena GheorghițăÎncă nu există evaluări
- Gametogeneza PDFDocument35 paginiGametogeneza PDFFlorinLaiosÎncă nu există evaluări
- Diviziunea CelularaDocument6 paginiDiviziunea CelularaElena TocÎncă nu există evaluări
- MeiozaDocument6 paginiMeiozaAlexia MariaÎncă nu există evaluări
- Diviziunea CelularaDocument9 paginiDiviziunea CelularaAlexander CrowlordÎncă nu există evaluări
- DiviziuneaDocument38 paginiDiviziuneaEugeniea RomanencoÎncă nu există evaluări
- DiviziuneDocument5 paginiDiviziuneIsvanca EmilianÎncă nu există evaluări
- New Microsoft Word Document 13Document3 paginiNew Microsoft Word Document 13cristiana.gabriela95Încă nu există evaluări
- Rezolvare Subiecte GeneticaDocument17 paginiRezolvare Subiecte GeneticaCristina IoanaÎncă nu există evaluări
- Diviziunea Celulara IndirectaDocument14 paginiDiviziunea Celulara IndirectaAna MariaÎncă nu există evaluări
- Subiecte Rezolvate ColocviuDocument18 paginiSubiecte Rezolvate ColocviuCristina IoanaÎncă nu există evaluări
- MEIOZADocument8 paginiMEIOZAAlexia MariaÎncă nu există evaluări
- Lucrare Practica La BiologieDocument3 paginiLucrare Practica La BiologieCristina SoloveiÎncă nu există evaluări
- Gametogeneza, Fecundatia, Spermatogeneza.Document17 paginiGametogeneza, Fecundatia, Spermatogeneza.Ana Lazar IvanÎncă nu există evaluări
- AdnDocument44 paginiAdnOxana GlibÎncă nu există evaluări
- Referat Tepurlui Norocel - Divizarea CelulelorDocument9 paginiReferat Tepurlui Norocel - Divizarea CelulelorPaula Tepurlui AmuzaÎncă nu există evaluări
- LP - 3 - Genetica Medicala - Anul Ii - MDDocument29 paginiLP - 3 - Genetica Medicala - Anul Ii - MDSendre CarmenÎncă nu există evaluări
- GametogenezaDocument9 paginiGametogenezaRoşu TeodoraÎncă nu există evaluări
- Mitoza MeiozaDocument2 paginiMitoza Meiozatusinean persidaÎncă nu există evaluări
- FIZIOLOGIEcurs 1Document5 paginiFIZIOLOGIEcurs 1Vladone VladonelolÎncă nu există evaluări
- Fiziologie Curs 1 BUNDocument4 paginiFiziologie Curs 1 BUNNicusor OnaciuÎncă nu există evaluări
- Fiziologie LP NR 5 Anul IDocument15 paginiFiziologie LP NR 5 Anul IAnna MariaÎncă nu există evaluări
- FIZIOLOGIE LP NR 4 Anul IDocument10 paginiFIZIOLOGIE LP NR 4 Anul IAnna MariaÎncă nu există evaluări
- Fiziologie LP NR 3 Anul IDocument13 paginiFiziologie LP NR 3 Anul IAnna MariaÎncă nu există evaluări
- Curs DZ2009 NouDocument142 paginiCurs DZ2009 NouLorena Stan100% (1)