Documente Academic
Documente Profesional
Documente Cultură
Fiziologia Analizatorul Vizual
Încărcat de
Ion CerlincaTitlu original
Drepturi de autor
Formate disponibile
Partajați acest document
Partajați sau inserați document
Vi se pare util acest document?
Este necorespunzător acest conținut?
Raportați acest documentDrepturi de autor:
Formate disponibile
Fiziologia Analizatorul Vizual
Încărcat de
Ion CerlincaDrepturi de autor:
Formate disponibile
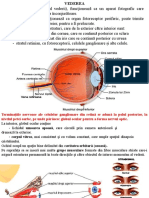
Fiziologia analizatorul vizual Vazul este cel mai important simt al omului si care furnizeaza aproape doua trei
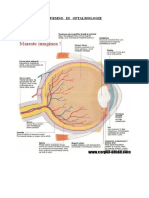
mi din toate informatiile prelucrate de creier. Cercetatorii sustin ca peste 80% din amintiri sunt inregistrate cu ajutorul ochilor. Receptorul periferic al vaz ului este ochiul (telereceptor) . In ciuda dimensiunii reduse, este un organ com plex: percepe formele, miscarile, reliefurile, culorile si diferentele de lumino zitate. Excitatia produsa de lumina este transmisa prin nervii si tracturile opt ice la scoarta cerebrala in aria 17 Brodmann, situata in jurul scizurii calcarin e, unde se transforma in senzatie vizuala. Ariile 18 si 19 situate in vecinatate , denumite ariile vizuale de asociatie, realizeaza perceptii mai vagi si servesc si ca arii asociative. Ochiul impreuna cu caile de transmitere si cu zonele de proiectie corticala formeaza aparatul sau analizatorul vizual. Ochiul are rolul de a furniza omului diverse informatii sub forma de imagini colorate, referitoar e la adancimea, distanta si miscarea obiectelor. Prin miscarea ochiului in toate directiile, putem vedea cea mai mare parte a mediului care ne inconjoara. Ochiu l se gaseste in cutia craniana, in orbita oculara, formata din tesut osos si tap etata cu tesut adipos pentru a proteja ochiul de lovituri. 1. Anatomia general a ochiului Prin structura sa, ochiul permite focalizarea lumi nii pe retin si transformarea semnalului luminos n semnale nervoase, care sunt tra nsmise apoi creierului. Ochiul omenesc poate sesiza semnale luminoase din domeni ul vizibil unde electromagnetice avnd lungimea de und cuprins ntre 400 nm (violet) s i 750 nm (rosu). Lumina reflectat de obiectele nconjurtoare determin modificri chimic e n celulele fotosensibile ale retinei, care transmit impulsuri nervoase n funcie d e forma, dimensiunile, poziia, orientarea i culoarea obiectelor. Globul ocular, or gan pereche, avand o form globular i are un diametru de cca. 2,5 cm. Este alcatuit dintr-un invelis, un aparat optic si un sistem receptor. Invelisul extern al och iului este alcatuit din sclerotica sau albul ochiului care in portiunea anterioa ra unde globul ocular devine mai bombat, devine transparenta si se numeste corne e si este avascular, bogat inervat, avnd o reea de terminaii nervoase libere sensibil e la durere, presiune, tact, cald i rece. Rolul corneii este protectie a irisului si pupilei. Corneea este primul mediu de refractie a luminii, numita si lentila optica. Sub sclerotica se afla un invelis colorat negru, puternic vascularizat, reprezentand tunica
mijlocie a ochiul cu functii nutritive, coroida. Prezint anterior irisul, cu un o rificiu numit pupila. Cele doua treimi posterioare ala coroidei sunt acoperite d e retina , care contine receptorii fotosensibili. Coroida se continua in partea anterioara cu o portiune ingrosata, corpul ciliar, format din fibre musculare ci rculare si longitudinale, pe care se insera o extremitate a ligamentului suspens or (ligamentul Zinn sau zonula) si o portiune inserate pe cristalin - disc trans parent care focalizeaza razele de lumina pe retina. Cristalinul este unul din me diile transparente ale ochiului. Acesta funcioneaz ca o lentil biconcava, razele de lumin captate suferind n ochi 3 refracii, dintre acestea 2 avnd loc n cristalin. Cri stalinul i modific forma n funcie de necesitate cu ajutorul muschilor din corpul cili ar. In fata cristalinului se afla irisul - disc musculos cu o gaura in mijloc, n umiata pupila. Rolul irisului este de a ajusta diametrul pupilei, regland astfel cantitatea de lumina care ajunge in cristalin. Irisul are pigmeni care dau culoa rea ochilor (albatri, cprui, verzi, negri). El delimiteaza pupila, format din fibr e radiale , care prin contractie produc midriaza( dilatatia pupilei) si circular e , care prin contractie provoaca mioza( contractie pupilara). Spatiul dintre cr istalin si cornee contine umoarea apoasa - lichid incolor, limpede , cu un conti nut sarac in proteine,ajuta la mentinerea curateniei corneei si indepartarea ger menilor. Este produs de ctre procesele ciliare n camera posterioar i ajunge n camera a nterioar prin pupil. Intre cristalin si retina (camera posterioara) se gaseste un alt lichid clar si gelatinos numit umoarea sticloasa sau corpul vitros. Cea mai importanta parte functionala a ochiului este retina care contine celule nervoase fotosensibile situate pe partea intern a peretelui globului ocular, care sunt responsabile de transformarea luminii n semnale nervoase. Acestea sunt de d oua tipuri: conuri sau bastonase cu structura asemanatoare, cele din urma avand o forma mai alungita, de unde deriva si denumirea. Ele sunt formate dintr-un seg ment extern, sub forma de cili modificati, care contin straturi transversale cu substante fotosensibile
proiectate in stratul extern pigmentar, dintr-un segment intern, bogat in mitoco ndrii si din regiunea nucleara, care se continua cu prelungiri ce fac sinapsa cu un strat de celule bipolare. Celulele nervoase bipolare transmit impulsul nervo s celulelor ganglionare, ai caror axoni alcatuiesc nervul optic, acesta parasest e ochiul prin discul optic, care nu contine nici conuri, nici bastonase, fiind d eci insensibil la stimulii luminosi, motiv pentru care se mai numeste si pata oa rba. Tot la acest nivel converg si vasele ochiului, care pot fi vizualizate si f olosite in diagnostic(astfel poate fi identificata arterioscleroza). Polul poste rior al ochiului este ocupat de pata galbena(macula lutea), in mijlocul careia s e afla foveea centralis, unde acuitatea vizuala este maxima. Este formata exclus iv din conuri. Conurile din aceasta regiune, spre deosebire de rest, fac sinapsa cu o singura celula bipolara si ganglionara, carora le corespunde in consecinta o singura fibra din nervul optic, ceea ce permite o analiza centrala mai exacta . In restul retinei, alaturi de conuri sunt prezente si bastonase, care predomin a numeric.
2. Studiul ochiului din punct de vedere geometric Ochiul poate fi echivalat cu o lentil convergent a crei distan focal este variabil. Co neea, membrana anterioar, transparent a ochiului, contribuie cu cea mai mare conve rgen la puterea optic a ochiului (puterea de focalizare a luminii pe retin). La ochi ul emetropimaginea unui obiect se formeaz clar pe retin indiferent de poziia obiectu lui fa de ochi. Acest lucru este posibil datorit modificrii convergenei cristalinului n procesul de acomodarela distan. In schimb, corneea contribuie cu o convergen const ant, dar mai mare dect a cristalinului (43 dioptrii fa de cele maxim 17 dioptrii ale cristalinului). Convergena ochiului este suma convergenelor corneei i cristalinulu i, deci este 60 dioptrii. Cu ct o lentil este mai convergent (are convergena mai mar e), cu att ea focalizeaz lumina mai aproape de lentil (are putere mai mare de focal izare). Cu ct lentila tinde s fie mai plat (crete raza de curbur a lentilei), cu att c onvergena sa scade. Corneea este mai bombat dect cristalinul, de aceea are converge na mai mare. Cristalinul realizeaz reglajul fin al focalizrii razelor de lumin pe re tin. Adaptarea la lumin In fata cristalinului se afla irisul, acesta reprezint o di afragm care limiteaz fluxul luminos ce cade pe retin i care micoreaz aberaiile cromati e i de sfericitate produse de lentilele ochiului. Irisul delimiteaza pupila, care poate varia intre 1,5-8 mm. Pupila este formata din celule netede, circulare si longitudinale. Contractia celor circulare produce mioza la iluminare excesiv ; a celor longitudinale, midriaza, cand luminozitatea este slaba. Mioza este produs a de stimuli parasimpatici, midriaza de stimuli simpatici. Mioza este produsa pr intr-un reflex cu cale aferenta prin nervii optici, cu centrii nervosi identici cu cei care produc si acomodarea cristalinului la distanta, calea efectoare fiin d aceiasi.
Atropina provoaca midriaza prin blocarea actiunii aceticolinei, prin contractia musculaturii radiale. Afectarea cai efectoare simpatice produce Sindromul Horner , constand in mioza de partea lezata, ca rezultat al preponderentei inervatiei p arasimpatice, prin caderea proapei superioare, din cauza paraliziei muschiului p alpebral superior ca si vasodilatatia zonei inervate de simpaticul lezat. Formar ea imaginii Imaginea este proiectata pe retina invers, intocmai cu aceea dintr-o camera fotografica. Este perceputa ca o imagine intr-o pozitie normala datorita interpretarii corticale. Razele de lumina strabat medii cu indice diferit de re fractie care converg la o distanta focala corespunzatoare retinei. Zonele de refractie sunt situate la nivelul contactului dintre aer si suprafata anterioara a corneei, dintre fata posterioara a acesteia si umoarea apoasa, dint re umoarea apoasa si suprafata anterioara a cristalinului si dintre suprafata lu i posterioara si umoarea sticloasa. Fiecare dintre aceste suprafete are un indic e de refractie diferit realizand in cele din urma 59 dioptrii, sistem optic in c are lentila este situata la 17mm de retina, distanta focala. O dioptrie este def inita ca puterea de refractie cu distanta focala de un metru. Daca aceasta dista nta este de m, puterea de refractie este egala cu 2 dioptrii, daca este de 10 cm cu 10 dioptrii. Pentru lentilele biconcave, dioptriile au valori negative si ex prima punctul la care se intalnesc razele divergente extrapolate de aceiasi part e cu obiectul. Puterea de refractie a ochiului se modifica astfel incat imaginea unui obiect aflat la distante diferite se formeaza pe retina. Pentru proiectia pe retina a unui obiect aflat la distanta, sistemul de refractie a ochiului este relaxat. Pentru ca imaginea unui obiect apropiat sa se formeze pe retina, crist alinul trebuie sa-si creasca convexitatea.
Puterea de refractie a cristalinului poate sa creasca la nou-nascut cu 17 dioptr ii. Cu timpul insa, prin pierderea elasticitatii cristalinului, puterea de acomo dare scade, ajungand pana la 2 dioptrii intre 40-50 ani. Acomodarea la distan Loca lizarea pe retina a imaginii in functie de distanta obiectului privit are loc pr in modificari adecvate a curburii cristalinului. Cristalinul este inserat intr-o capsula elastica ce are tendinta de a-l mentine, ca si elasticitatea lui propri e, sub forma bombata, apropiata de sfera. Cristalinul este insa inconjurat de li gamentul suspensor, care se afla in tensiune si il turteste. Cand obiectul privi t se apropie de ochi tensiunea ligamentului suspensor scade, datorita contractie i muschiului ciliar, format din fibre meridionale si circulare. Fibrele circular e realizeaza un sfincter. In ochiul emetrop neacomodat (relaxat, privete la infin it) muchiul ciliar este relaxat. Tensiunea exercitat de ligamentele suspensoare de termin ntinderea (aplatizarea) cristalinului;raza sa de curbur este maxim, iar conve rgena minim. Imaginea se formeaz clar pe retin. In cazul n care convergena cristalinul i nu s-ar modifica, imaginea unui obiect mai apropiat de ochi s-ar forma n spatel e retinei (aceasta se ntmpl la ochiul presbit). De aceea, cnd obiectul se apropie de ochi, cristalinul trebuie s i mreasc puterea de convergen pentru a strnge razele tot retin, formnd imaginea clar. Cum se realizeaz acest lucru? Prin contracie, muchiul ci liar scade tensiunea n ligamentele suspensoare, cristalinul tinde s revin la forma sa natural i se bombeaz; raza sa de curbur scade, deci convergena crete. Aceasta este acomodarea la distan. Pentru a se putea acomoda, ochiul trebuie s aib un cristalin e lastic. Odat cu naintarea n vrst, cristalinul devine mai rigid (presbitism), de aceea se folosesc lentile convergente de corecie, care mresc convergena ochiului. Lumina venit de la un obiect ndeprtat sau apropiat este focalizat n acelai punct prin curbarea cristalinului: Vederea clar se realizeaz ntre dou puncte: punctum proximum (pp) i punctum remotum (p r). Pp este punctul cel mai apropiat care poate fi vzut clar cu acomodare maxim. P r este punctul cel mai deprtat, vzut clar fr acomodare. La ochiul normal (emetrop) p p = 25 cm, pr la aproximativ 6 metri (infinit oftalmologic). Lumina venita de la un obiect apropiat sau departat este focalizat in acelasi pu nct prin curbarea cristalinului.
3. Defectele geometrice ale vederii Defectele geometrice ale vederii (ametropiile) sunt: - defecte axiale (se refer l a dimensiunile globului ocular) - defecte de curbur (forma dioptrilor) - defecte de indice (indicii de refracie ai mediilor transparente) - defecte de elasticitat e (proprietile mecanice ale cristalinului) Miopia Miopia axial este cea mai frecven t form de miopie. Axul anteroposterior al globului ocular este mai lung, imaginea se formeaz naintea retinei. Pp i pr se afl mai aproape de ochi. Miopia axial se corec teaz cu lentile divergente. In miopia de curbur cristalinul are o curbur mai mare d ect la ochiul normal i acest lucru duce la mrirea convergenei (de obicei, acest tip de miopie este legat de oboseal). In miopia de indice, care apare n stri patologice , crete indicele de refracie datorit creterii concentraiei saline. Hipermetropia In ochiul hipermetrop imaginea se formeaz n spatele retinei, pp se a fl mai departe dect la ochiul emetrop. Hipermetropia axial apare cnd axul anteropost erior al globului ocular este mai scurt dect n mod normal. Hipermetropia de curbur este caracterizat printr-un cristalin mai alungit. Cristalinul trebuie s se bombez e n permanen pentru a aduce imaginea pe retin. Corectarea hipermetropiei se face cu lentile convergente.
Presbitismul Este o ametropie de elasticitate care apare, n general, dup vrsta de 4 0 de ani. Bombarea se face mai dificil. Se folosesc lentile convergente pentru a vedea obiectele apropiate. Astigmatismul Astigmatismul este o ametropie de curb ur. Razele de curbur ale dioptrilor nu sunt egale de la un meridian la altul al di optrilor (mai ales pentru cornee). Forma dioptrilor nu mai este sferic. Corectare a se face cu ajutorul lentilelor cilindrice. 4.Mecanismul fotoreceptor 4.1. Structura cristalinului, corneei si retinei Cristalinul este compus din ap i proteine transparente (care nu absorb lumina vizibil). Proteinele sunt dispuse or donat, n cca. 20000 de straturi subiri, concentrice, al cror indice de refracie crete dinspre periferie spre centru. Aceast stuctur permite trecerea luminii cu pierder i minime, ceea ce determin aspectul transparent al cristalinului. In plus, crista linul (ca i corneea) acioneaz ca un filtru care absoarbe o mare parte din radiaiile electromagnetice cu lungimi de und mici (ultraviolete, X, g), avnd rol n protecia reti nei. Cristalinul este ncapsulat n sacul capsular (cristaloida), care este susinut d e fibrele zonulare (zonulele lui Zinn). Cristalinul este o lentil biconvex asimetr ic, avnd razele de curbur de 10 mm i respectiv 6 mm n stare neacomodat. Corneea este u n esut avascular i transparent, a crui grosime variaz ntre 0,5 mm (la
centru) i 1,2 mm (la periferie). Este format din 5 straturi. Dinspre exterior spre interior acestea sunt: - epiteliul corneal format din celule cu putere mare de regenerare n prezena oxigenului (lentilele de contact inhib regenerarea); acest str at este meninut umed prin lichidul lacrimal; - stratul Bowman este foarte reziste nt, conine fibre de colagen; protejeaz stroma; - stroma corneal este un strat gros, conine n jur de 200 de straturi orizontale de fibre de colagen dispuse ordonat; d e aceea undele electromagnetice mprtiate de diferitele fibre interfer distructiv, as tfel nct lumina incident trece practic fr pierderi n fibrele de colagen, iar corneea a pare transparent; stroma conine de asemenea foarte puine cheratocite; practic, stro ma corneal nu are capacitate de regenerare; - membrana posterioar este foarte subir e, acelular; are rol de susinere a endoteliului; - endoteliul corneal este un mono strat celular aflat n contact cu umoarea apoas; regleaz fluxurile de fluide i solvii n tre umoarea apoas i stroma corneal; de asemenea mpiedic hidratarea corneei (aceasta a r duce la vedere nceoat). Retina este un strat senzorial care conine celule sensibile la lumin (celulele cu conuri i celulele cu bastonae) i alte celule nervoase care pr eiau, transmit i proceseaz semnalele produse de celulele fotosensibile. Datorit pro ceselor evolutive, retina prezint un aranjament inversat: pe direcia luminii, celule le senzoriale se afl n spatele stratului de fibre nervoase. In retin sunt prezente 6 tipuri de celule, dispuse n straturi succesive: Celulele epiteliului pigmentar alctuiesc stratul distal format dintr-un monostrat de celule epiteliale, care au o form regulat, asemntoare unor prisme hexagonale, cu seciune hexagonal n planul retin i. Aceste celule epiteliale conin un pigment, melanina, care absoarbe puternic lu mina, evitndu-se astfel procesele de mprtiere a luminii prin reflexii interne n ochi (melanina din celulele coroidei contribuie de asemenea la minimizarea reflexiilo r; de aceea, n cazul albinismului, absena melaninei determin o slab acuitate vizual). Poriunea intern,puternic pigmentat, a celulelor epiteliale pigmentare formeaz numer oase procese filiforme care se extind printre bastonaele i conurile celulelor foto receptoare, avnd ca rol principal fagocitoza segmentului extern al celulelor foto receptoare i adugarea de material membranar nou la segmentul intern al acestora. S tratul urmtor este cel al celulelor fotoreceptoare, celulele cu conuri i celulele cu bastonae; acestea sunt neuroni specializai care conin pigmeni fotosensibili. Celu lele fotoreceptoare produc semnale nervoase pe care le transmit prin eliberare d e neurotransmitori. Cantitatea de neurotransmitori eliberai depinde de numrul de foton i recepionai. Semnalele nervoase sunt apoi procesate de ali neuroni ai retinei, i an ume: celulele orizontale, bipolare, amacrine i ganglionare.
Celulele fotoreceptoare sunt orientate cu extremitatea fotosensibil nspre coroid, f iind n contact direct cu epiteliul pigmentar. Repartiia lor n retin nu este uniform. In fovee se afl numai celule cu conuri, n timp ce densitatea celulelor cu bastonae crete nspre periferie. In pata oarb celulele fotoreceptoare lipsesc complet.
Urmeaz stratul de celule orizontale, care fac sinaps cu celulele fotoreceptoare (6 -50 celule fotoreceptoare), realiznd conexiuni ntre celulele fotoreceptoare i celul ele bipolare. Pe lng transmiterea semnalului nervos, celulele orizontale au un rol foarte important n asigurarea unui contrast foarte bun al imaginii. Acesta se re alizeaz prin procesul de inhibiie lateral, n care celulele orizontale pot inhiba sel ectiv transmisia nervoas pe cile de comunicare care nu pornesc din imediata vecinta te a senzorilor puternic luminai. Astfel, atunci cnd lumina cade pe retin, receptor ii din zona de inciden sunt excitai puternic, n timp ce receptorii aflai n jurul zonei de inciden sunt mai puin luminai. Celulele orizontale elimin semnalele senzorilor sl ab luminai, crescnd n acest fel contrastul imaginii i acuitatea vizual. Intr-un strat succesiv sunt dispuse celulele bipolare, neuroni vizuali (se mai numesc i neuron i bipolari) care realizeaz legturi ntre celule fotoreceptoare sau orizontale i celul e amacrine sau ganglionare. O celul bipolar poate primi semnale de la o celul fotor eceptoare fie n mod direct, fie prin intermediul unui celule orizontale. In zona foveal corespondena este biunivoc: fiecare con realizeaz legturi sinaptice cu o bipol ar i fiecare bipolar cu o ganglionar. Fiecare ganglionar primete astfel informaii de l un singur con. Spre periferia foveei i n afara acesteia, mai multe celule recepto are realizeaz conexiuni sinaptice cu o bipolar i mai multe bipolare trimit informaii unei singure ganglionare. Celulele bipolare amplific procesul de inhibiie lateral. Exist cca. 10 tipuri diferite de celule bipolare care primesc semnale
de la celule cu conuri i un singur tip de celule bipolare care realizeaz legturi cu celule cu bastonae. Celulele amacrine mediaz semnalele nervoase ntre celulele bipo lare i alte celule amacrine sau ganglionare. Ca i celulele orizontale, celulele am acrine acioneaz lateral, afectnd semnalizarea celulelor bipolare nvecinate, dar prez int un grad de specializare mult mai pronunat. Exist cca. 40 tipuri diferite de cel ule amacrine, majoritatea fiind lipsite de axon. Fiecare tip celular amacrin se conecteaz cu anumite tipuri de neuroni bipolari i genereaz un anumit tip de neurotr ansmitori. Celulele amacrine intervin n procesarea imaginii n special la nivelul reg lrii luminozitii imaginii i al deteciei micrii obiectelor.
Ultimul strat este format din celulele ganglionare. Acestea fac sinaps cu cele bi polare, iar axonii lor alctuiesc nervul optic. Pata oarb (papila cieca), lipsit de celule fotoreceptoare,este locul n care nervul optic se ndreapt spre corpii genicul ai laterali, dup ce strbate nveliul globului ocular. Pata oarb apare ca un oval alb, a vnd o arie de cca. 3 mm2. In exteriorul globului ocular, axonii celulelor ganglio nare sunt mielinizai. In medie, o celul ganglionar poate primi semnale produse de c ca. 100 de celule fotoreceptoare, dar acest numr variaz mult cu localizarea pe ret in. La om, retina conine n jur de 1,2 1,5 milioane celule ganglionare. Exist cel puin 5 clase de celule ganglionare, dintre care trei sunt specializate n procesarea i nformaiilor vizuale n funcie de micare (clasa W), culoare (clasa X) i intensitatea lu minii (clasa Y). Celelalte dou clase cuprind celule ganglionare care asigur reflex ul pupilar (adaptarea la lumin) i meninerea ritmurilor circadiene; o mare parte din aceste celule sunt fotosensibile conin un fotopigment, melanopsina, care le perm ite s rspund direct la lumin, chiar n absena celulelor cu conuri i bastonae. Dintre n onii retinieni, numai celulele ganglionare produc poteniale de aciune de tip tot s au nimic, acestea fiind transmise creierului prin nervul optic. Ceilali neuroni r etinieni produc poteniale electrice gradate i transmit semnalul nervos modulnd cant itatea eliberat de neurotransmitori n funcie de fluxurile membranare de sarcini elect rice, care variaz n mod continuu (conducie electrotonic). In acest fel,
intensitatea semnalului primar, generat de celulele fotoreceptoare, este proporio nal cu cantitatea de lumin recepionat. Retina are o grosime de 0,4 mm. In fovee, o mic zon de depresiune avnd un diametru de 0,3 mm, retina este mai subire: celulele s enzoriale au dimensiuni minime i sunt dispuse compact pe prile laterale, ntr-un aran jament de mozaic hexagonal, pentru a nu obtura lumina i pentru a obine o densitate maxim de fotoreceptori. Aici exist numai celule cu conuri. Foveea (fovea centrali s) este zona responsabil pentru vederea central i percepia vizual a detaliilor cu mar e acuratee; de asemenea determin n mare parte percepia culorilor. Foveea se afl n cent rul maculei, o zon circular galben, aflat n apropiere de centrul retinei, cu un diame tru de cca. 1,5 mm. Pigmentaia galben a maculei se datoreaz unor carotenoizi (n spec ial zeaxantina i luteina) prezeni n terminaiile nervoase ale celulelor cu conuri din aceast zon, avnd rol n protecia celulelor fotosensibile ale maculei fa de radiaiile X i g, pe care aceti pigmeni le absorb. Macula (macula lutea sau pata galben) are o d ensitate mare de celule cu conuri i este specializat pentru a asigura o bun acuitat e vizual; este de asemenea responsabil pentru vederea central. Dei macula ocup mai pui n de 0,01% din cmpul vizual, cca. 10% din axonii nervului optic preiau informaiile transmise de celulele fotoreceptoare din zona foveal. Fa de alte zone ale retinei, foveea produce informaie vizual cu cea mai mare acuratee. S-a determinat recent c r etina transmite informaii ctre creier cu o vitez de 8,75 megabit/s. Dei foarte mare, aceast vitez este limitat de consumul de energie metabolic necesar pentru desfurarea proceselor fiziologice care sunt implicate n procesele recepiei vizuale. La verteb rate, n timpul dezvoltrii embrionare retina i nervul optic apar ca prelungiri ale c reierului n dezvoltare, de aceea retina este o parte a sistemului nervos central (SNC). De altfel, singura parte a SNC care poate fi vizualizat n mod direct este r etina. Stratul de celule senzoriale ale retinei (retina senzorial) este n contact cu stratul epitelial pigmentar, dar nu este fixat de acesta dect n zona de emergen a nervului optic, denumit discoptic, pat oarb sau papil. De aceea, dac se produce o mi c ruptur a retinei, fluidul care intr prin fisur va ndeprta retina de epiteliu, determ innd desprinderea progresiv a retinei. In acest caz retina nu mai poate prelua flu xurile de nutrieni dinspre coroid, ceea ce duce la moartea (lent) a retinei. Netrat at, o desprindere de retinei duce n 6-8 luni la pierderea ireversibil a vederii.
4.2. Structura si functia celulelor fotoreceptoare
Celulele fotoreceptoare realizeaz funcia de traducere a semnalului vizual radiaia e lectromagnetic din domeniul vizibil n semnal electric. Celula cu bastona este alctui t din dou pri: segmentul extern (SEB), care are o form alungit, cilindric, de bastona segmentul intern (SIB). Segmentul extern este
fotoreceptorul propriu-zis, iar cel intern conine nucleul i organitele celulare, a vnd rol metabolic. Cele dou segmente sunt conectate printr-un cil. Terminaia sinapt ic a SIB formeaz sinaps cu o alt celul nervoas, de tip bipolar sau orizontal. Celulele receptoare cu bastona, avnd o mare sensibilitate la lumin (o singur celul poate sesi za un singur foton), sunt responsabile pentru vederea scotopic, la luminozitate s czut, fr distingerea culorilor (vedere acromatic, alb-negru). Retina uman conine cca. 00 milioane de celule cu bastona. SEB are o structur special, coninnd un mare numr de discuri membranare (pn la 2000) suprapuse, care mresc astfel suprafaa de interacie cu lumina. Membrana discurilor este format din subuniti membranare (cca. 5 nm diametr u) n centrul crora se gsete pigmentul fotosensibil rodopsina (107-108 molecule/basto na). Rodopsina, protein trans-membranar care traverseaz membrana de 7 ori ( helixuri), este alctuit din opsin (partea proteic) i cromoforul retinal (aldehida vitaminei A, partea prostetic); cromofor = grupare care absoarbe lumina. Membrana bastonaului c onine numeroase canale de Na+ care sunt deschise de cGMP. La ntuneric concentraia c elular de cGMP este mare, astfel nct exist un influx pasiv de Na+ i Ca2+ (Ca2+ n propo rie de 10-15%) prin aceste canale de Na+ (curent de ntuneric). Datorit acestui cure nt, la ntuneric membrana este depolarizat, avnd un potenial de membran n jur de -40 mV , mai mare dect potenialul de repaus de -65 mV al celulelor nervoase. La aceste va lori ale potenialului de membran, canalele de Ca2+ din membrana plasmatic se deschi d, conducnd la creterea concentraiei de Ca2+ n citoplasm. Concentraiile de Na+ i Ca2+ n continuare la valori constante datorit pompelor ionice Na+ -K+ din SIB i mecanis mului antiport 3 Na+ /1 Ca2+ din SEB, care mpiedic acumularea excesiv a cationilor n citoplasm. Concentraia mare de Ca2+ intracelular determin exocitoza veziculelor ca re conin neurotransmitori, n special glutamat. Moleculele de glutamat sunt eliberate n spaiul sinaptic i se leag la receptorii de glutamat din terminalul celulei nervoa se postsinaptice; ele pot inhiba anumii neuroni bipolari i pot excita ali neuroni b ipolari, n funcie de tipul de receptori pe care acetia i prezint. Legarea glutamatulu i la receptori declaneaz rspunsul celular (o variaie a potenialului de membran, care s e propag spre cellalt capt al celulei, unde se declaneaz o nou eliberare de neurotrans mitori), asigurnd transmiterea semnalului nervos la urmtoarele celule nervoase. La l umin, n urma fotoexcitrii i activrii rodopsinei, concentraia de cGMP scade, canalele d e Na+ se nchid, curentul de ntuneric dispare i membrana se hiperpolarizeaz. Potenialu l celular poate ajunge la -80 mV, depinznd de intensitatea luminii. Canalele de C a2+ se nchid, ca urmare celula elibereaz mai puin glutamat dect la ntuneric. Variaia d e potenial membranar produs n neuronii bipolari depinde att de tipul neuronului bipo lar, ct i de cantitatea de glutamat eliberat de celulele
fotoreceptoare, deci de cantitatea de lumin recepionat. Legturile complexe interneur onale i tipurile diferite de neuroni retinieni aparinnd aceleiai clase contribuie la procesarea complex a informaiei vizuale la nivelul retinei, nainte ca aceasta s fie preluat de nervul optic. Semnalul nervos excit n final neuronul ganglionar, care g enereaz trenuri de poteniale de aciune de tip tot sau nimic. Acestea, pe calea nerv ului optic, ajung n corpii geniculai i apoi n scoara cerebral (scizura calcarin) unde roduc senzaia vizual. Bastonaele au o sensibilitate foarte mare: un singur foton po ate duce la blocarea intrrii n celul a 106 sarcini pozitive amplificare de putere. Fotonul este doar trigger (declanator), restul se datoreaz energiei proceselor met abolice. Celulele fotoreceptoare nu se divid, dar materialul membranar este renno it continuu prin procese de fagocitoz ntreinute de celulele epiteliului pigmentar. Celulele cu conuri permit perceperea culorilor n condiii de luminozitate accentuat (vedere fotopic diurn). Retina uman conine n jur de 5-6 milioane celule cu conuri. El e sunt activate n condiii de iluminare puternic au un prag ridicat de activare. Au forma de con, iar n loc de discuri au o membran faldurat. Neurotransmitorul eliberat de celulele cu conuri este acetilcolina. Pigmentul fotosensibil al conurilor est e iodopsina, care are o structur similar rodopsinei. In retina uman s-au identifica t trei tipuri de conuri, cu sensibilitate cromatic diferit i care conin trei tipuri de pigmeni iodopsinici: eritrolab (cu maximul de absorbie la = 564 nm), clorolab ( -5 34 nm) i cianolab ( - 420 nm). Cei trei pigmeni difer prin civa aminoacizi n opsin ce determin absorbia diferit a luminii. Faptul c nu sesizm dect lumina vizibil se dato eaz faptului c radiaiile electromagnetice cu lungimi de und <400 nm nu ajung pe reti n, acestea fiind absorbite de cornee, cristalin i pigmenii maculei. Anumiipacieni la care a fost ndeprtat cristalinul au raportat capacitatea de a observa n ultraviolet . Culorile albastru( l=450-492nm), verde (l=492-575nm) si rosu ( l=647-723nm) sunt den umite primare, pentru ca din amestecul lor poate fi realizata orice alta culoare , inclusiv cea alba. Pentru verde, culoarea complementara este cea rosie si inve rs. Acest fapt poate fi demonstrat si pe plan de senzatie, culoarea complementar a aparand dupa inlaturarea celei stimulatoare. O teorie veche, teoria tricolora sau tricromatica a lui Young-Helmholtz, astazi acceptata ca teoria de baza insa corectata si completata, sustine ca exista trei tipuri de conuri, cate unu pentr u fiecare din cele trei culori primare si ca fiecare produce un curent de actiun e de o anumita frecventa. Codul de transmisie a culorilor este diferit de cel pr esupus de Young-Helmholtz.
Anomalii de percepere a culorii Exista diferite deficiente de percepere a culori i, care merg pana la orbirea cromatica, in sensul ca imaginea este perceputa doa r in alb-negru-cenusiu. Anomaliile de perceptie se clasifica pe baza incapacitat ii de a percepe cele trei culori principale, anomalie semnificand o slabire, iar anopie o orbire cromatica. Culorile la care se refera aceste particularitati su nt desemnate prin prefixul prot-(rosu) , deuter-(verde) si trit(albastru). Pot f i afectate una sau toate cele trei receptivitati cromatice. Daltonismul este inc apacitatea de a percepe culoarea rosie( protanopsie) sau verde (deuteranopsie) s au ambele.
Ciclul biochimic al rodopsinei Cromoforul rodopsinei, retinalul, se afl la ntuneri c n configuraia 11-cis. Prin fotoactivare, retinalul trece n configuraia all-trans i se desprinde de opsin. Transformarea direct mpreun cu transformarea invers reprezint c iclul Wald. In starea n care retinalul s-a desprins, rodopsina devine rodopsin act ivat R* i interacioneaz cu traductina T (macromolecul proteic membranar din clasa prot inelor G), care la rndul ei este activat. Traductina, care n stare neactivat are leg at o molecul de guanozin difosfat (GDP), va forma un complex R*T-GDP care pierde G DP i leag GTP (guanozin trifosfatul) devenind R*T-GTP. Acest complex este instabil i se descompune n R* i T-GTP. R* se combin din nou cu traductina neactivat etc. R* s e poate combina cu pn la 500 molecule de T. Complexul T-GTP se numete traductin acti vat. Aceasta activeaz apoi o molecul de fosfodiesteraz (PDE); se formeaz complexul PD E-T-GTP (fosfodiesteraz activat)
care acioneaz asupra c-GMP (acid guanozin-monofosforic ciclic). c-GMP este transfo rmat n acid guanozin-monofosforic 5'-GMP. Scderea concentraiei de c-GMP determin nchi derea canalelor de Na+. Fosfodiesteraza activat pierde o grupare fosfat i devine P DE-T-GDP care la rndul su se descompune n TGDP i PDE. Acestea din urm reintr n cicluri e biochimice respective. Se pot distinge deci urmtoarele cicluri: ciclul Wald (RR*), ciclul traductinei, ciclul PDE i ciclul c-GMP (c-GMP este refcut sub aciunea g uanilat ciclazei, care se activeaz la scderea concentraiei de Ca2+) Prin aceste cic luri se produce o amplificare considerabil a semnalului. Astfel, o molecul de rodo psin activeaz 50 T/s i o traductin transform 2000 4000 cGMP/s. Ca rezultat final, un singur foton poate induce hidroliza a cca. 100000 c-GMP/s. Aceasta duce la nchide rea unui mare numr de canale de Na+; ionii de Na+ i Ca2+ se acumuleaz n exteriorul S EB i determin hiperpolarizarea membranei, proporional cu intensitatea stimulului lu minos. Printr-o serie de alte procese biochimice este limitat mecanismul de ampl ificare i, la ntuneric, R* devine din nou inactiv R (rodopsin kinaza catalizeaz scder ea afinitii R* pentru T, prin fosforilare).
Tehnici de investigare si tratament Exist o serie de tehnici pentru diagnosticare a diferitelor afeciuni ale retinei, cum ar fi: - oftalmoscopia pentru investigare a retinei; - electroretinografia se msoar activitatea electric a retinei; - tomogra fia cu coeren optic genereaz o tomogram a retinei, cu mare rezoluie spaial; permite alizarea detaliilor histologice.
Imaginea retinei observat cu un oftalmoscop. Pigmentaia galben a maculei nu este vi zibil ntr-o astfel de imagine. 5. Notiuni de patologie ale globului ocular 1. Retinopatia diabetica Diabetul poate induce cu mare probabilitate pierderea v ederii. Diabetul provoaca o reducere progresiva a circulatiei capilarelor in ret ina. Ischemia produsa determina dezvoltarea unor noi vase fragile de sange in re tina. La concentratii mari de glucoza sau fructoza in sange, aceste vase se pot rupe, producand hemoragii in umoarea vitroasa. Laserul este utilizat pentru foto coagularea vaselor rupte de sange sau pentru tratarea ischemie retinale. Efectul este cresterea raportului de oxigen in retina ceea ce determina regresia vaselo r de sange. 2. Glaucomul Umoarea apoas este secretat n camera posterioar de corpul ciliar, trece prin spaiul ngust dintre cristalin i iris, apoi prin pupil ajunge n camera anterioar. De aici este drenat n sistemul venos, prin reeaua trabecular situat n apropierea baze i irisului. Presiunea exercitat de umoarea apoas menine forma curbat a corneei.
Glaucomul este caracterizat de creterea presiunii intraoculare, datorat fie unei s ecreii excesive de umoare apoas, fie obturrii fluxului n reeaua trabecular. Glaucomul netratat determin degenerarea progresiv a nervului optic prin distrugerea celulelo r ganglionare, ducnd n timp la pierderea ireversibil a vederii. 3. Cataracta
In cataract, cristalinul devine opac, blocnd sau mprtiind o parte din lumina incident. Imaginile apar estompate. Cauzele cataractei nu sunt bine cunoscute n prezent, d ar diabetul, glaucomul, expunerea la radiaii (n radioterapie), expunerea prelungit la soare (la radiaii ultraviolete), precum i diferite medicamente (de exemplu, tam oxifen, amiodoron, sau utilizarea de lung durat a aspirinei) sunt unii dintre fact orii de risc asociai cataractei. Netratat, cataracta duce la pierderea vederii. Ce l mai larg utilizat tratament al cataractei, facoemulsificarea, folosete o sond cu ultrasunete pentru a fragmenta (emulsifica) cristalinul, care este apoi nlocuit. I n anumite cazuri se utilizeaz laserul pentru emulsificarea cristalinului, dar las erul nu este la fel de puternic precum ultrasunetele i nu poate fragmenta cristal inul n cazurile avansate. Dup emulsificare, fragmentele cristalinului sunt absorbi te i eliminate din sacul capsular. Este implantat apoi un cristalin artificial pe baz de silicon sau material acrilic. Pentru implant se folosete un injector de mi ci dimensiuni, care este introdus printr-o incizie practicat n cornee. Materialul injectat se nfoar i se ancoreaz spontan n spatele pupilei, n sacul capsular rmas lib p emulsificare, realiznduse n acest fel un implant de cristalin foldabil.
BIBLIOGRAFIE 1. 2. 3. 4. Note de curs Biofizica, Adrian Iftimie si Constanta Gan ea Fiziologie, Petru Groza http://www.scritube.com/biologie/Vederea203217818.php http://altamedicina.com/afectiuni-oculare/ Nota: sursele imaginilor folosite: O mare parte din imagini sunt produse de Cate dra de Biofizica Medicala a UMF Carol Davila Bucuresti; ele pot fi folosite libe r,
pentru scopri didactice sau educationale (CreativeCommons Lic.). Lista completa acestor imagini poate fi gasita la sediul catedrei. Restul imaginilor sunt folos ite ca citari din: - Netter Atlas of Human Anatomy, C.G. Edition - Principles of Neural Science, E. Kandel, Ed 4. - ICO's Principles and Guidelines of a Curricu lum for Ophthalmic Education of Medical Students, www.icoph.org/med/ - Cursul de optica biologica al "Physics Department of Texas University" - Imagini sub lice nta C.C. preluate de la wikipedia.com - Lectii interactive de Biologie, ed. 1, m anual interactiv produs de Intuitext.ro
S-ar putea să vă placă și
- Fiziologia Analizatorul VizualDocument20 paginiFiziologia Analizatorul VizualAlecsandra Tache100% (1)
- Analiza Si Fiziologia Analizatorului VizualDocument6 paginiAnaliza Si Fiziologia Analizatorului VizualGuță NicoletaÎncă nu există evaluări
- Fiziologia Analizatorul VizualDocument10 paginiFiziologia Analizatorul VizualGiani Daniel GuțăÎncă nu există evaluări
- Ochiul Si Defectele de VedereDocument13 paginiOchiul Si Defectele de Vedereloredanaiacob100% (4)
- OCHIUL Si URECHEADocument2 paginiOCHIUL Si URECHEAIoana BobeicăÎncă nu există evaluări
- Lucrare Conjunctivita GonococicaDocument86 paginiLucrare Conjunctivita GonococicaAna Maria Maxim100% (2)
- Anatomia OchiuluiDocument6 paginiAnatomia OchiuluiCristina AurelÎncă nu există evaluări
- Proiect Absolvire An 2Document66 paginiProiect Absolvire An 2Mariana Daminescu100% (3)
- C7 Analizatorii Ochiul UrecheaDocument8 paginiC7 Analizatorii Ochiul UrecheaAlina FlorentinaÎncă nu există evaluări
- Analizatorul Vizual-BiologieDocument13 paginiAnalizatorul Vizual-BiologieMocanuÎncă nu există evaluări
- L9. Analizatorul VizualDocument4 paginiL9. Analizatorul VizualLaura FloreaÎncă nu există evaluări
- Analizatorul VizualDocument20 paginiAnalizatorul VizualOana Stefania Pintilii100% (1)
- OCHIULDocument7 paginiOCHIULIonica Madalina100% (1)
- Temă-Analizator VizualDocument3 paginiTemă-Analizator VizualHăpăianu ElisaÎncă nu există evaluări
- Analizatori 2Document78 paginiAnalizatori 2Ioana- Cristina ChituÎncă nu există evaluări
- Analizatorul VizualDocument11 paginiAnalizatorul VizualMiruna Deleanu MariaÎncă nu există evaluări
- Analizatorul VizualDocument41 paginiAnalizatorul Vizualmariana77100% (2)
- Sistemul Senzorial Optic Vederea OchiulDocument9 paginiSistemul Senzorial Optic Vederea OchiulIulia Inglis0% (1)
- Formarea Imaginii Pe RetinaDocument7 paginiFormarea Imaginii Pe RetinaRosalinda MironÎncă nu există evaluări
- Fiziologia Analizatorului VizualDocument18 paginiFiziologia Analizatorului VizualDieFerÎncă nu există evaluări
- Analizatorul VizualDocument9 paginiAnalizatorul VizualAlessandra McaÎncă nu există evaluări
- Misca Bristena-Igiena Analizatorului VizualDocument21 paginiMisca Bristena-Igiena Analizatorului VizualInu PriescuÎncă nu există evaluări
- 05 Curs Receptie VizualaDocument17 pagini05 Curs Receptie VizualaLoveMed09Încă nu există evaluări
- Fiziologia OchiDocument6 paginiFiziologia OchiDorobantu MarianaÎncă nu există evaluări
- Untitled DocumentDocument43 paginiUntitled Documentsculpt05Încă nu există evaluări
- Elemente de Optica BiologicaDocument20 paginiElemente de Optica BiologicaMurariu MadalinaÎncă nu există evaluări
- Organele de Simt Ale MamiferelorDocument9 paginiOrganele de Simt Ale MamiferelorRadu Ionut Alex100% (1)
- Cap. 12 Organele de SimțDocument7 paginiCap. 12 Organele de SimțDragos BirzuÎncă nu există evaluări
- Analizatorii Sunt Sisteme Morfofunctionale Prin Intermediul CaroraDocument11 paginiAnalizatorii Sunt Sisteme Morfofunctionale Prin Intermediul Caroraisidor_mihaelaÎncă nu există evaluări
- Suport de Curs - Anatomia Analizatorului VizualDocument19 paginiSuport de Curs - Anatomia Analizatorului VizualClaudia Serban100% (1)
- OCHIULDocument14 paginiOCHIULAndreiÎncă nu există evaluări
- Analizatorul Vizual La OmDocument15 paginiAnalizatorul Vizual La OmCotaru LuciaÎncă nu există evaluări
- Organele de SimțDocument31 paginiOrganele de SimțPaul vlad 06Încă nu există evaluări
- Anatomia OchiuluiDocument1 paginăAnatomia OchiuluiAndreea Si AtatÎncă nu există evaluări
- VazulDocument2 paginiVazultiovidiuÎncă nu există evaluări
- Licență GlaucomulDocument22 paginiLicență GlaucomulAlex CmnÎncă nu există evaluări
- Analizatorul Vizual FiziologieDocument6 paginiAnalizatorul Vizual FiziologieNinaPodarascuÎncă nu există evaluări
- Elementele Ochiului SuntDocument6 paginiElementele Ochiului Suntbbertha13Încă nu există evaluări
- Ochiul UmanDocument4 paginiOchiul UmanMolnarRaduÎncă nu există evaluări
- Proiect OptometrieDocument24 paginiProiect OptometrieIonela-Georgiana Urse100% (7)
- Anatomia OchiuluiDocument2 paginiAnatomia OchiuluiMihaela CanțărÎncă nu există evaluări
- Analizatorul VizualDocument4 paginiAnalizatorul VizualSorin GabrielÎncă nu există evaluări
- OchiulDocument4 paginiOchiulNicolae ElenaÎncă nu există evaluări
- Analizatorul Vizual OCHIULDocument9 paginiAnalizatorul Vizual OCHIULAna Maria ToporascuÎncă nu există evaluări
- Analizatorul Vizual.Document28 paginiAnalizatorul Vizual.Vali Catalin100% (1)
- Referat OchiulDocument5 paginiReferat OchiulAna-Maria TrandafirescuÎncă nu există evaluări
- OftalmologieDocument21 paginiOftalmologiecraciun monica100% (1)
- Referat Biologie OchiulDocument4 paginiReferat Biologie OchiulCiocarlan Mihai25% (4)
- AnatomieDocument28 paginiAnatomieEduard TomițaÎncă nu există evaluări
- Anatomia Analizatorului VizualDocument28 paginiAnatomia Analizatorului VizualDaniela Maria GaneaÎncă nu există evaluări
- Optica Elementara - 2021Document45 paginiOptica Elementara - 2021IoanaJohannaaÎncă nu există evaluări
- Analizatorul VizualDocument7 paginiAnalizatorul VizualDaniela MariaÎncă nu există evaluări
- NEUROPSIHOLOGIE: elementele de bază ale problemeiDe la EverandNEUROPSIHOLOGIE: elementele de bază ale problemeiEvaluare: 5 din 5 stele5/5 (1)
- Ai grija de viata, de tine, de ceilalti si de naturaDe la EverandAi grija de viata, de tine, de ceilalti si de naturaÎncă nu există evaluări
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 4 din 5 stele4/5 (8)
- Mysterium Coniunctionis. Cercetări asupra separării şi unirii contrastelor sufleteşti în alchimie. Volum suplimentar. Aurora consurgensDe la EverandMysterium Coniunctionis. Cercetări asupra separării şi unirii contrastelor sufleteşti în alchimie. Volum suplimentar. Aurora consurgensEvaluare: 5 din 5 stele5/5 (1)