Documente Academic
Documente Profesional
Documente Cultură
Sistemul Muscular
Încărcat de
TamaraBmBTitlu original
Drepturi de autor
Formate disponibile
Partajați acest document
Partajați sau inserați document
Vi se pare util acest document?
Este necorespunzător acest conținut?
Raportați acest documentDrepturi de autor:
Formate disponibile
Sistemul Muscular
Încărcat de
TamaraBmBDrepturi de autor:
Formate disponibile
HISTOLOGIE GENERAL CORNELIA AMLINEI
ESUTUL MUSCULAR
esutul muscular reprezint un esut nalt specializat, cu proprietate esenial
contractibilitatea. n vederea ndeplinirii acestei funcii celulele musculare conin filamente
contractile, n principal Actina i Miozina. Structura acestora genereaz fora necesar contraciei
musculare.
Celulele musculare prezint origine mezodermic, diferenierea lor realizndu-se printr-un
proces gradual de alungire, cu sinteza simultan de elemente miofibrilare.
Celulele musculare sunt denumite i cu termenul impropriu de fibre, datorit formei lor
alungite, totui ele trebuie difereniate de fibrele adevrate, componente ale esutului conjunctiv.
esutul muscular poate fi clasificat funcional n:
Voluntar: esutul muscular striat scheletal
Involuntar: esutul muscular cardiac i neted visceral.
Clasificarea morfologic, bazat pe aspectul citoplasmei celulelor musculare, este
urmtoarea:
esut muscular striat: scheletal i cardiac
esut muscular neted: visceral.
Organitele celulare sunt denumite diferit fa de corespondentele lor din alte celule, datorit
aspectului esutului muscular comparat cu carnea de pete (sarcos, n limba greac), dup cum
urmeaz:
Citoplasma (fr miofibrile) reprezint sarcoplasma
REN reprezint Reticolul Sarcoplasmic (RS)
membrana celular sau plasmalema reprezint sarcolema.
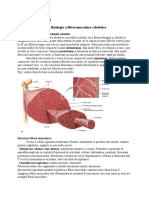
esutul muscular striat scheletal
Localizare
esutul muscular striat scheletal se gsete att la nivelul musculaturii asociate scheletului,
ct i la nivelul buzelor, limbii i poriunii superioare a esofagului.
Aspect n microscopia optic
n microscopia optic se constat prezena de mnunchiuri de fibre musculare lungi (pn la
30 cm), de form cilindric n inciden secional longitudinal sau rotund-ovalare sau poligonale
n seciunile transversale sau oblice (diametrul de 10-100 m). Se constat aspectul acidofil al
citoplasmei, cu striaii longitudinale determinate de prezena miofibrilelor (colonetele Leydig); n
incidena transversal se constat un aspect intat determinat tot de prezena miofibrilelor, aspect
denumit cmpurile Cohnheim. Celulele musculare striate scheletale sunt multinucleate, fapt
datorat fuzionrii celulelor embrionare mononucleate-mioblaste. Nucleii sunt situai sub
sarcoplasm, de form alungit, n seciunile longitudinale i mai rotunjit, n incidenele
transversale. Dispoziia periferic i multitudinea nucleilor faciliteaz diagnosticul diferenial cu
fibrele musculare cardiace i netede care prezint nuclei unici, centrali.
La microscoapele optice cu o rezoluie nalt se constat alternana de discuri clare,
izotrope, monorefringente (I), cu discuri ntunecate, anizotrope, birefringente (A).
Discul I, cu o lungime de 0,8 m este compus din 2 hemidiscuri, fiind bisectat de o band
ntunecat Z (stria Amici sau Zwishensheibe).
Discul A, cu lungimea de 1,5 m, prezint o treime foarte ntunecat, birefringent n
lumina polarizat, o treime central mai puin ntunecat, monorefringent- banda Hensen / Helle
1
HISTOLOGIE GENERAL CORNELIA AMLINEI
(H) care conine central o band subire mai colorat, linia M i nc o treime ntunecat,
birefringent.
Spaiul delimitat de dou benzi Z, coninnd un hemidisc I, un disc A i nc un hemidisc I,
constituie unitatea morfo-funcional a celulei musculare, sarcomerul.
Aspect n microscopia electronic i ultrastructur
Microscopia electronic a evideniat c pe fiecare band Z, pe ambele fee, se inser
filamente subiri de Actin, fr a se ntlni cap la cap.
Linia Z are aspect de linie frnt datorit emiterii a cte 4 piciorue unite prin - actinin,
nebulin i filamin. n jur se gsesc filamente intermediare de desmin (scheletin), vimentin,
ancorarea la sarcoplasm realizndu-se prin filamente de vinculin, talin, prin formarea de
costamere.
Printre filamentele subiri se gsesc filamente mai groase de Miozin cu ambele extremiti
libere. Ele ptrund pe o anumit distan printre filamentele subiri, sunt unite la mijloc prin
intermediul unor puni proteice de miomezin, o protein-enzim (creatin-kinaz), proteina C i
titina (conectina).
Se pot concluziona urmtoarele:
discul I conine doar filamente subiri-monorefringent
zona A conine dou tipuri de filamente care determin birefringena
zona H conine numai filamente groase
linia M reprezint puntea proteic.
Miofibrila reprezint o suit repetitiv de sarcomere identice ca ultrastructur.
n seciunile transversale pe la nivelul discului I se observ 6 filamente de Actin dispuse n
unghiurile unui hexagon.
Seciunile pe la nivelul discului A prezint 6 filamente de Actin dispuse n hexagon cu un
filament de Miozin central.
Filamentele de Miozin sunt dispuse n unghiurile unui triunghi i la nivelul liniei M
capetele triunghiului sunt unite prin puni proteice.
Raportul total dintre filamentele de Miozin i cele de Actin este de .
Citoscheletul
Citoscheletul este alctuit din.
filamente intermediare necontractile care stabilizeaz structura descris: desmin
(scheletina), vimentin, sinemin
circa 20 de proteine accesorii, cum ar fi: -actinina, filamin, amorfin, proteina C,
miomezina, nebulina, vinculina, talina, titina (conectina), alturi de distrofin, dispus de
jur mprejurul miofibrilei i sincrofin situat intern tot cu dispoziie concentric.
Distrofia muscular Duchenne este o boal genetic caracterizat prin imposibilitatea
sintezei de distrofin, manifestndu-se prin degenerarea celulelor musculare striate scheletale,
uneori i a celor cardiace i nlocuirea prin esut fibro-adipos, urmat de deces pn la vrsta de 20
de ani.
Tipuri de fibre musculare striate
n raport cu proporia dintre miofibrile/sarcoplasm exist trei tipuri de fibre musculare
striate:
Tipul 1, roie, lent, de postur, n care predomin sarcoplasma bogat n mioglobin, care
cupleaz oxigenul n mod similar Hemoglobinei, mitocondrii bogate n citocrom, numeroase
enzime oxidative, ATP-az redus, mai puine miofibrile. Acest tip de fibr este mai ngust,
fibrele fiind subiri i determin o contracie puternic, susinut, lent, repetitiv, avnd o mare
rezerv de oxigen i energie generat prin fosforilarea oxidativ.
Tipul 2B, alb, rapid, n care predomin miofibrilele, mioglobina fiind mai redus,
sarcoplasma i mitocondriile mai slab reprezentate, enzimele oxidative sunt mai puine, ATPaze, fosforilaze abundente; formeaz fibre mai mari, cu o contracie foarte rapid, foarte fin,
de durat mai scurt, mai redus ca amplitudine, pe baza glicolizei anaerobe.
2
HISTOLOGIE GENERAL CORNELIA AMLINEI
Tipul intermediar, 2A, cu un coninut intermediar de mioglobin, mitocondrii i coninut
enzimatic, cu o contracie rapid, prelungit, pe baza fosforilrii oxidative i a glicolizei
anaerobe.
La om, n general, exist toate cele trei tipuri, cu unele predominene. Apariia acestor tipuri
de fibre st sub dependena inervaiei motorii care este n raport cu funcia muchiului; un neuron
nu inervez concomitent i fibre albe i fibre roii.
Sistemul T-RS
Sistemul T este reprezentat de tubi transveri, aprui prin invaginri ale sarcolemei la
fiecare jonciune disc I - Disc A. Sarcomerul are doi tubi T, specializri necesare pentru ca unda de
depolarizare s ajung la miofibrile. Dup ce ptrund n interiorul celulei se ramific foarte mult,
astfel nct fiecare miofibril va avea cte un tub T la jonciunea disc I - Disc A.
Sistemul RS este compus din RS i este dispus pe fiecare segment dintre doi tubi T. n
vecintatea fiecrui tub T, de-o parte i de alta exist o cistern terminal, deci sistemul RS
reprezint un spaiu nchis.
Seciunea transversal la nivelul unui tub T va prezenta de-o parte i de alta cte o cistern
terminal realizndu-se astfel triade. Fiecare sarcomer prezint dou triade, ntre elementele triadei
apar din loc n loc zone mai electronodense- piciorue joncionale. RS conine 2 proteine: ATPaza Ca2+-Mg2+-dependent i calsechestrina, avid de Calciu.
Organizarea molecular a filamentelor contractile
Filamentele subiri:
Actina este alctuit din Actina globular G care se organizeaz ntr-un lan polarizat
dublu-spiralat Actina fibroas F. Actina F are o lungime de 1 m i un diametru de 7
nm i prezint o zon cu afinitate pentru Miozin- situs activ. Pe Actina F se gsesc alte
filamente, dup cum urmeaz:
Tropomiozina (Tm) este o molecul fibroas, linear, cu o lungime de 41 nm, alctuit
din dou lanuri polipeptidice (dimer). O molecul de Tropomiozin acoper 7 molecule
de Actin G, fiind dispus n depresiunea spiralei Actinei F, moleculele vecine cuplnduse ntr-o manier cap-coad.
Troponina (Tn) este o protein globular dispus ntre moleculele de Tropomiozin,
fiind alctuit din 3 subuniti (trimer):
Tn t, la nivelul cozii, se cupleaz la tropomiozin
Tn c se cupleaz la Calciu, prin 4 situsuri
Tn i este inhibitorie pentru interaciunea Actin- Miozin.
Tropomodulina (Tm) se inser la capetele filamentelor de Actin, realiznd, prin
blocarea polimerizrii, stabilizarea lungimii acesteia.
Filamentele groase: Miozina este o molecul proteic mare, cu o lungime de 1,5 m i un
diametru de 15 nm, cu o form spaial asemntoare unei crose cu dou capete.
Tija este alctuit din dou lanuri polipeptidice helicoidale (lanuri grele) i capul este
alctuit din dou perechi de lanuri uoare, realiznd un segment globular bilobat. Cele dou
poriuni pot fi scindate prin tratarea cu Papain.
n alctuirea filamentelor sunt 250-300 de molecule cu tijele paralele. Capetele proemin la
exterior doar n zona n care filamentele groase se ntreptrund cu cele subiri (zona H este
lipsit de capete globuloase), avnd o dispoziie n spiral.
La nivelul extremitii globuloase se gsete o zon cu mare afinitate pentru Actin i cu
activitate ATP-azic.
Proteinele asociate Miozinei sunt: miomezina care realizeaz puni transversale cu
activitate creatin-kinazic, titina (conectina) care ancoreaz Miozina la linia Z i proteina C
care se cupleaz la Miozin n vecintatea liniilor M.
Tripsina scindeaz Miozina n:
meromiozin grea (H), care corespunde capetelor globuloase i gtului
3
HISTOLOGIE GENERAL CORNELIA AMLINEI
meromiozina uoar (L) care corespunde restului tijei.
Zona de jonciune prezint o mare flexibilitate, putnd realiza unghiuri de pn la 120,
flexibilitate care face posibil mecanismul contraciei musculare prin glisare.
Transmiterea impulsului de contracie
Membrana plasmatic este polarizat pozitiv la exterior datorit ionilor Na i negativ la
interior datorit ionilor K. Depolarizarea membranei presinaptice determin deschiderea canalelor
de Calciu (Ca) cu intrarea acestuia n axon, ceea ce determin eliberarea Acetilcolinei din butonii
terminali, gsirea receptorilor specifici i inducerea depolarizrii membranei ntr-un punct
(Potenial de aciune), fenomen urmat de extensia rapid pe toat sarcolema. Depolarizarea
membranei o face refractar la alt stimul.
ntre butoni i pliuri exist o cantitate mare de Acetilcolinesteraz care hidrolizeaz
Acetilcolina, ceea ce conduce la repolarizarea i receptivitatea pentru noi stimuli. Acetilcolina este
reciclat, colina este returnat terminaiei axonale pentru a fi recombinat cu acetilcoenzima A din
mitocondrii i a fi stocat n vezicolele sinaptice. Membranele vezicolelor sinaptice golite sunt
reciclate prin vezicole endocitozice tapetate de clatrin.
n intoxicaiile cu pesticide se inactiveaz acetilcolinesteraza, aprnd contracii musculare
prelungite.
n Miastenia gravis, se produc Autoanticorpi cu o organizare spaial asemntoare
Acetilcolinei, determinnd blocarea receptorilor specifici.
Depolarizarea ajunge n vecintatea miofibrilelor prin sistemul tubilor T, se produce
activarea ATP-azei din RS, cu deschiderea canalelor de Ca din membrana RS. Acest fenomen
va determina eliberarea Ca de pe calsechestrin, nivelul su crescnd brusc la nivelul miofibrilei.
Nivelul Ca este responsabil pentru declanarea modificrilor biochimice responsabile de contracia
muscular.
Intensitatea contraciei unui muchi depinde de numrul fibrelor musculare care intr n
contracie.
O fibr nervoas motorie mpreun cu totalitatea fibrelor musculare striate inervate de
aceasta formeaz o unitate motorie (neuromotorie). Toate celulele musculare dintr-o unitate se
contract la unison, dup legea totul sau nimic. n cazul unui numr redus de fibre albe
contraciile musculare sunt de finee iar n cazul unui numr ridicat de fibre roii, contracia este
puternic. Fora contraciei unui muchi este proporional cu numrul unitilor motorii care intr
n aciune.
Inervaia motorie menine troficitatea fibrelor musculare inervate. n poliomielit, datorit
degenerrii fibrelor nervoase, se produce hipotrofia muscular.
Mecanismul contraciei
n timpul contraciei se produce o scurtare a sarcomerelor de la 20 pn la 50%. Mecanismul
este cel de glisare bazat pe o secven de asociere i disociere a moleculelor de Actin i Miozin.
La nivelul fibrelor musculare n repaus, ATP-ul este fixat pe captul globulos al Miozinei, nu
este hidrolizat de ATP-aza miozinic deoarece complexul Tn-Tm se interpune ntre cele dou zone
de afinitate reciproc Actin- Miozin.
Cnd Ca este pompat de RS la nivelul miofibrilelor se realizeaz demascarea zonei de
afinitate Actin-Miozin prin urmtoarele secvene: Ca se fixeaz pe Tn c, formndu-se un
complex Ca-Tn; acest complex determin o alt orientare spaial a complexului Tn-Tm
(Tropomiozin); deplasarea Tm demasc poriunea de Actin capabil s activeze ATP-aza, se
produce hidroliza ATP-ului rezultnd ADP i energie care mic braul Miozinei (ca o balama),
determinnd deplasarea filamentelor de Actin.
Astfel, datorit contraciei, discul A rmne neschimbat, discul I se ngusteaz,
filamentele subiri ptrunznd pe o distan mai mare printre filamentele groase, pn la dispariia
zonei H.
Prin hidroliza ATP-ului se decupleaz Actina de Miozin i ia sfrit un ciclu. ntr-un ciclu
sunt activate cteva capete ale Miozinei i n urmtoarele cicluri alte capete sunt puse n micare,
4
HISTOLOGIE GENERAL CORNELIA AMLINEI
permind celor care s-au contractat s-i refac ATP-ul. Astfel, prin pai succesivi, Actina este
tras printre filamentele de Miozin.
Cnd Ca dispare, Tn c pierde cuplarea cu acesta i se produce refacerea orientrii Tn, Tm,
oprirea hidrolizei ATP ducnd la relaxare.
Condiiile necesare relaxrii musculare sunt:
s nu existe Ca (pompat din citosol n RS pentru a fi cuplat la calsechestrin)
s existe ATP pe captul Miozinei
s nu fie demascat zona Actinei cu afinitate pentru Miozin care acioneaz ca un
co-factor al ATP-azei.
Condiiile necesare contraciei musculare sunt urmtoarele:
aflux de Ca
demascarea Actinei (co-factor prin formarea complexelor Ca- Tn)
activarea ATP-azei cu degajare de energie.
Se poate constata c ATP-ul este necesar att contraciei ct i relaxrii musculare. Lipsa sa
explic apariia rigiditii cadaverice (rigor mortis) care apare post-mortem datorit meninerii
cuplajului Miozin- Actin, pentru c nu se mai reface ATP-ul.
ATP-ul este produs de enzimele mitocondriale prin -oxidarea acizilor grai i din Glucoz,
pe calea ciclului Krebs (glicoliz mai rapid). Refacerea ATP-ului se realizeaz din fosfocreatin
( la nivel mitocondrial prin utilizarea glicogenului i a acizilor grai), care, prin fosforilare, se
transform n fosfat i acesta n ADP.
Prin glicoliz aerob dintr-o molecul de Glucoz se produc 38 molecule de ATP.
Prin glicoliz anaerob, dintr-o molecul de Glucoz se genereaz dou molecule de ATP, cu
producerea de lactai, piruvai care determin oboseala i febra muscular.
Placa motorie
Placa motorie este reprezentat de o jonciune neuromuscular (sinaps
neuromuscular). Fibra nervoas motorie se ramific ntr-o serie de mici extensii butonate care
conin neuromediatorul Acetilcolin stocat n vezicole sinaptice, REN, mitocondrii. Fibra nervoas
pierde n aceast regiune teaca de mielin rmnnd nconjurat doar de teaca celulelor Schwann.
Astfel se realizeaz membrana presinaptic.
Fanta sinaptic conine un material asemntor lamei bazale amorfe.
Membrana postsinaptic se prezint astfel: sarcolema pierde membrana bazal, la nivelul
jonciunii, formeaz pliuri joncionale (fante sinaptice II) pe care sau n structura crora se gsesc
molecule-receptori specifici pentru acetilcolin, mitocondrii, ribozomi, RER.
n botulism (toxinfecie alimentar cauzat de Clostridium botulinum) este inhibat
eliberarea de acetilcolin la nivelul jonciunii mio-neuronale, boala manifestndu-se prin paralizie
muscular, tulburri vizuale i fiind fatal dac nu este tratat.
Fusul neuro-muscular
Fusul neuro-muscular realizeaz reglajul intrinsec al contraciei musculare, funcionnd ca
un mecano-receptor n principal pentru forele de ntindere. Acesta are un aspect fuziform, cu
lungimea de 1-3 mm, fiind nconjurat de fibre musculare scheletale obinuite (fibre extra-fusale).
La exterior prezint un nveli subire de esut conjunctiv. n interiorul su se gsesc 8-10 fibre
musculare striate (fibre intrafusale) cu anumite particulariti:
fibre cu poriunea central mai dilatat, cu 3-4 nuclei (fibre cu nuclei n sac); la extremiti
se afl discuri I i A
fibre cilindrice, care conin la extremiti discuri I, A i la mijloc o multitudine de nuclei (fibre
cu nuclei n lan sau n irag).
n spaiul periaxial se gsete un coninut de consisten fluid.
Celulele primesc terminaii nervoase senzoriale de dou tipuri:
I, mai groase, cu o conducere rapid, n raport cu ambele tipuri de fibre
II, mai subiri, cu o conducere lent, n raport cu cu celulele cu nuclei n lan (terminaii
nervoase n ieder).
5
HISTOLOGIE GENERAL CORNELIA AMLINEI
Gradul de contracie al unui muchi este recepionat prin gradul de contracie al acestor fibre
striate specializate, transmis prin nervii senzoriali la mduva spinrii, reglnd rata (rspunsul
fazic) i durata (rspunsul tonic) ntinderii musculare.
Fibrele de tip I fac sinaps cu neuronul motor de la acelai nivel al mduvei spinrii iar fibrele
de tip II fac sinaps i prin intermediul neuronilor asociativi. Astfel, n mod reflex, este comandat
contracia muscular necesar meninerii posturii.
Fibrele specializate prezint i inervaie motorie (fibre ). Acestea realizeaz plci neuromusculare n zonele n care exist striaii. Suprastimularea muscular determin stimularea
fibrelor eferente care determin contracia cu contrabalansarea ntinderii.
Un organ asemntor se gsete i n apropierea jonciunilor musculo-tendinoase, printre
fibrele tendinoase, avnd rolul de reglare a posturii-Organul Golgi. Acesta este alctuit din fibre de
colagen ncapsulate, nconjurate de ramificaii terminale ale nervilor senzitivi tip 1B. Contraciile
prea puternice determin creterea tensiunii n tendon, stimularea fibrelor senzitive 1B, avnd
rezultat inhibarea fibrelor motorii . Reflexul rotulian reprezint un exemplu al prezenei acestor
organe Golgi.
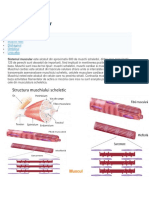
Organizarea general a muchilor striai scheletali
Fibrele musculare striate din cadrul unui muchi se organizeaz n fascicole, cu fibrele
paralele ntre ele, orientate n lungul muchiului. Fiecare fibr conine n jurul sarcolemei un nveli
de tipul membranei bazale (MB). Printre fibre se gsesc mici septuri de esut conjunctiv lax (a
crui fibre reticulare intr n alctuirea MB), cu capilare sanguine i fascicole nervoase, formnd
endomisiumul.
Fiecare fascicol este investit cu un sept conjunctiv mai bogat n fibre, coninnd arteriole,
venule, limfatice, nervi, formnd perimisiumul.
nveliul muchiului este constituit din epimisium sau aponevroz (cnd musculatura este
etalat). Epimisiumul este alctuit din esut conjunctiv dens semiordonat, perforat de nervi i de
vase care se ramific n perimisium i endomisium.
La extremiti se gsesc structuri conjunctive care ataeaz muchiul pe elementul ce
urmeaz a fi mobilizat (os, cartilagiu), formnd tendoanele. Jonciunea muchi-tendon reprezint o
jonciune de tip contiguitate, realizat prin jonciuni aderente la nivelul benzilor Z, cu pliuri i
ngrori datorit occludinelor i desmogleinelor. Structurile conjunctive ale muchiului i ale
tendonului sunt n relaie de continuitate, dup cum urmeaz:
Endomisium- endotenoniu
Perimisium- peritenoniu
Epimisium- epitenoniu.
Celulele satelite reprezint celule regenerative, situate n lamina extern a celulei musculare
striate scheletale. Acestea genereaz prin difereniere i fuzionare fibre musculare scheletale.
Histogeneza musculturii striate
Miotomul mezodermic reprezint originea mioblatilor care sunt mononucleari, fuziformi.
Acetia vor migra n vecintatea pieselor cartilaginoase, vor suferi diviziuni succesive, sintetizeaz
Actina, apoi Miozina, care se vor structura n filamente. Se vor profila astfel miofibrilele dispuse
periferic. Datorit interveniei unor citokine se va produce sinciializarea, cu realizarea unor celule
multinucleate cu nuclei centrali, cu continua producere de miofibrile, miofilamente, cptnd falsa
denumire de Miotom. Sinteza va continua, cu dislocarea nucleilor i poziionarea lor spre periferie,
zona central fiind ocupat de miofibrile, formndu-se fibra matur. Aproximativ 5% din mioblati
rmn blocai n maturare, fiind celulele satelite.
Modificri adaptative/patologice
Hipertrofia nseamn creterea dimensiunilor fiecrei fibre, sub influene de natur
nervoas, cu creterea numrului filamentelor i cu antrenarea unor celule satelite.
Hipotrofia semnific micorarea datorit lipsei stimulilor nervoi motori, cum ar fi posttraumatic.
Hiperplazia nseamn mrirea numrului fibrelor musculare.
6
HISTOLOGIE GENERAL CORNELIA AMLINEI
esutul muscular cardiac
Localizare
esutul muscular cardiac reprezint componenta principal a inimii, miocardul, fiind situat
i la emergena vaselor mari, n tunica lor medie.
Histogenez
n timpul dezvoltrii embrionare, din mezoblastul precordal apar celule care se aliniaz
cordonal, fr a fuziona i realizeaz complexe joncionale intercelulare. Diferenierea ulterioar se
realizeaz printr-un proces treptat de alungire i ramificare, cu sinteza concomitent de proteine
miofibrilare.
Organizare general
esutul muscular cardiac este organizat sub form de reea, cu multiple anastomoze, reea
format din fibre musculare cardiace care determin apariia a numeroase ochiuri pline cu esut
conjunctiv. Aceast structur este adaptat la necesitatea de contracie a cavitilor cardiace.
Celulele musculare cardiace sunt asamblate n coloane de cte 2-4 celule (miocite).
Sarcolema prezint un strat de tipul laminei bazale pe care se inser fibre reticulare din
esutul conjunctiv interstiial al miocardului.
La exteriorul sarcolemei se gsete un esut conjunctiv lax denumit endomisium.
esutul conjunctiv dispus n ochiurile reelei de coloane musculare reprezint
corespondentul perimisiumului. Acesta este un esut conjunctiv lax, bogat n vase sanguine,
limfatice, nervi i fibre elastice (mai ales n pereii atriali), n continuitate cu esutul conjunctiv al
endocardului, epicardului, cu scheletul fibros al inimii i cu pereii vaselor mari. Vasele sanguine
realizeaz un plex capilar (aproximativ un capilar la 3 celule cardiace). Fibrele nervoase simpatice
i parasimpatice realizeaz plexuri nervoase miocardice.
Se constat o heterogenitate a fibrelor musculare cardiace, care determin delimitarea a
trei tipuri de miocite cardiace, dup cum urmeaz:
celulele musculare cardiace contractile, comune sau efectoare care se gsesc la nivel atrial, ct
i la nivel ventricular
celulele musculare cardiace ale sistemului de conducere (excito-conductor, cardionector,
embrionar) care sunt grupate sub forma nodului sino-atrial, nodului atro-ventricular,
fascicolului atrio-ventricular His cu ramurile sale i reelei Purkinje.
celulele musculare cardiace cu proprieti secretorii (endocrine, mioendocrine) care se
gsesc mai ales n atriul drept.
Celula muscular cardiac contractil prezint o form cilindric, capetele fiind bifide
sau, mai rar trifide, cu o lungime de 85-100 m i un diametru de 15-20 m. Sunt mononucleate,
uneori binucleate, cu nucleii situai central, alungii, cu o lungime de aproximativ 8,3 m,
hipocromi, cu nucleol vizibil.
Sarcoplasma este acidofil i prezint o dubl striaie:
Striaia longitudinal (colonetele Leydig) este conferit de agregrile miofibrilelor, mai subiri
i cu dispoziie mai neregulat dect n fibra muscular striat scheletal. Miofibrilele ocolesc
nucleul, astfel nct delimiteaz o arie perinuclear.
Striaia transversal este evident i se datoreaz alternanei de discuri clare i ntunecate,
vizibile n microscopia optic i n microscopia n contrast de faz, n mod similar fibrelor
musculare striate scheletale.
Seciunile transversale n MO prezint un contur rotunjit, sau mai poligonal, unele fibre
fiind mai neregulate i mai alungite datorit ramificrilor. Nucleii sunt dispui central, au aspect
rotunjit n cazul incidenelor secionale transversale. Miofibrilele apar ca mici zone acidofile care
HISTOLOGIE GENERAL CORNELIA AMLINEI
confer sarcoplasmei un aspect intat-cmpuri Cohnheim, reprezentnd de fapt mai ales grupri
artefactuale de miofibrile.
Sarcomerul este mai scurt dect cel al fibrei musculare striate scheletale, cu o lungime de
aproximativ 2,5 m, ocazional putnd ajunge la 7-8 m.
Filamentele subiri sunt compuse din:
Actin
Tropomiozin
Troponin
Tropomodulin.
Filamentele groase sunt reprezentate de Miozin.
Discul intercalar sau stria scalariform sau stria Eberth reprezint o caracteristic a
fibrei musculare cardiace, sub forma unei linii transversale dispuse la interfaa dintre dou celule
musculare cardiace vecine, de form linear sau n trepte, datorit lungimii inegale a miofibrilelor.
Aceast structur poate fi vizualizat n preparatele uzuale dar mai ales cu Hematoxilin-acid
fosfotungstic (Hematoxilin feric). n cazul n care stria prezint un aspect de treapt, i se descriu
dou regiuni:
componenta transversal, n unghi drept fa de miofibrile, care este nsoit de interdigitarea
fibrelor musculare adiacente
componenta lateral, paralel cu miofibrilele, neted.
Jonciunile intercelulare de la nivelul striei scalariforme sunt de trei tipuri:
Fasciile aderente constituie poriunea principal a componentei transverse, mai ntinse dect
maculele, lsnd un spaiu de 15-20 nm ntre cele dou membrane plasmatice; ME evideniaz
un material electronodens n acest spaiu. Se nsoesc de condensarea citoplasmei adiacente.
Srvesc drept situsuri de ancorare a filamentelor subiri de Actin din sarcomerul terminal pe
membrana plasmatic, astfel nct reprezint n esen hemidiscuri Z.
Maculele aderente (desmozomi) se gsesc i n poriunile transversale i n cele laterale. Rolul
desmozomilor este de a mpiedica disocierea celulelor, n momentul contraciei.
Jonciunile gap (nexus) constituie elementul structural major al componentei laterale. Acest tip
de jonciune permite trecerea n ambele direcii a moleculelor cu greutate molecular mic, fr
ca acestea s difuzeze n spaiul intercelular. Realizeaz o comunicare ionic i contribuie la
sincronismul funcional al fibrelor musculare cardiace.
Sarcoplasma este mai abundent perinuclear, coninnd mai mult RER, complex Golgi i
incluzii de lipofuscin (telolizozomi, pigment de uzur) sub forma unor granule galben-brune,
vizualizate optim n coloraia Hematoxilin feric (metoda Heidenheim).
n ceea ce privete raportul dintre organitele din sarcoplasm, fibra muscular cardiac se
aseamn cu fibra muscular striat scheletal tip 1: predomin mioglobina i mitocondriile
(sarcozomii) fa de miofibrile. Sarcozomii ocup cel puin 40% din volumul sarcoplasmei (fa de
doar 2% n muchiul scheletal), ceea ce reflect necesitatea continu de metabolism aerob. Pe lng
mitocondriile juxtanucleare, se gsesc i mitocondrii mari, avnd creste dense, dispuse n mod
condensat printre miofibrile, extinse pe ntreaga lungime a sarcomerelor.
Incluziile de glicogen sunt condensate printre miofibrile. n condiii de stress, o mic parte
din glicogen poate fi scindat n glucoz i utilizat ca surs de energie. n mod obinuit ns sursa
energetic este reprezentat de acizii grai, transportai ca lipoproteine ctre muchiul cardiac i
depozitai ca trigliceride n numeroase picturi lipidice.
Sistemul T-RS
Tubii T, ca invaginri ale sarcolemei, nu se gsesc la jonciunea dintre discurile A i I, ca n
celulele musculare striate scheletale, ci la nivelul benzilor Z, fiind mai largi, tapetai de un strat
glicoproteic continuu cu stratul care acoper sarcolema. Un sarcomer are numai un tub T,
asemnndu-se astfel cu muchiul striat al batracienelor.
Sistemul RS prezint cte o cistern terminal n vecintatea tubilor T i altele
subsarcolemale. Prezint piciorue joncionale sarcoplasmice, ca prelungiri emise de ctre
8
HISTOLOGIE GENERAL CORNELIA AMLINEI
cisterna terminal, apropiindu-se foarte mult de tubul T i coninnd canale de Ca ++ dependente de
ATP-az. Sistemul longitudinal este mai puin dezvoltat i anastomozat la nivelul zonei mijlocii
(zona H). RS conine calsechestrin care leag Ca++ i ATP-az care elibereaz Ca++ .
Din loc n loc apar diade, alctuite dintr-un tub T i o cistern RS, n locul triadelor (dou
cisterne RS i un tub T), ntlnite la muchiul striat.
Mecanismul contraciei
Contracia miocardului se realizez n special pe baza degradrii aerobe a glucozei i a
acizilor grai i prin fosforilarea oxidativ.
Mecanismul contraciei se realizeaz prin glisarea filamentelor subiri printre cele groase.
n stare de repaus, ADP-ul i fosfatul anorganic rmn cuplate la ATP-aza din captul
globular al moleculei de miozin.
Influxul de Ca++ se realizeaz prin pompe ionice de Ca++ care devin active ca urmare a
stimulilor vegetativi ce activeaz Adenilat-ciclaza membranar cu producerea de acid adenilic-ciclic
responsabil de deschiderea canalelor de Ca++. Acest aport extra-celular de Ca este necesar deoarece
RS este mai puin dezvoltat i rezerva intra-celular de Ca++ este mai redus la fibra muscular
cardiac fa de cea scheletal. Influxul de Ca++ determin o secven de evenimente similar celei
existente la fibra muscular striat scheletal: cuplarea Ca++ la Tn c, schimbarea configuraiei Tn
i modificarea poziiei Tm. Aceasta determin expunerea situsului Actinei G de cuplare la
Miozin. Cuplarea Miozinei la Actin conduce la eliberarea ADP i fosfatului iar captul globulos
al Miozinei se ndreapt ctre zona H. Actina va urma aceeai direcie fiind cuplat la Miozin.
Glisarea captului Miozinei reexpune situsul de cuplare al ATP, ATP se cupleaz la ATP-az,
elibernd astfel capul Miozinei de Actin. ATP este scindat i Miozina revine la configuraia
iniial.
Diferene fenotipice
Celulele musculare cardiace contractile prezint diferene dup localizarea lor:
celulele cardiace atriale prezint dimensiuni mai mici, tubi T mai redui numeric
celulele cardiace ventriculare prezint tubi T mai largi.
Aspecte patologice
Hipertrofia celulelor cardiace contractile poate apare la sportivi, la marii butori de bere, n
cazul unor diverse obstacole (stenoze/insuficiene valvulare), n Boala Hipertensiv, diametrul
fibrelor putnd ajunge pn la 30 m. Hipertrofia, nefiind nsoit de creterea numrului sau a
diametrului capilarelor, determin apariia unei insuficiene n irigaie, rezultnd dilatarea i
insuficiena cardiac.
n cazul ocluziei arterelor coronare care irig cordul, prin trombui sau coaguli, (adesea
eliberai de pe o plac de aterom) sau datorit unor spasme vasculare prelungite se produce
moartea celular n segmentul de miocard corespunztor (necroz)-Infarct Miocardic. n cazul
unor Infarcte miocardice nefatale, esutul necrotic este reparat prin nlocuirea cu esut fibros,
deoarece nu exist celule satelite dup prima copilrie, deci lipsete capacitatea de regenerare.
Inervaia
Fibrele adrenergice realizeaz sinapse din loc n loc (en passant) stabilind o relaie de
apropiere cu celulele musculare cardiace, rmnnd un mic spaiu ntre acestea (100-200 nm).
Fibrele senzoriale sunt responsabile de perceperea durerii miocardice.
Celulele musculare cardiace ale sistemului de conducere
Celulele musculare cardiace ale sistemului de conducere fac posibil contracia ritmic,
spontan i sincron a miocardului, fr stimuli direci de la sistemul nervos. Dup criterii morfofuncionale se deosebesc urmtoarele tipuri de celule aparinnd sistemului excitoconductor:
Celulele situate n Nodul Sino-Atrial (Keith-Flack) au o funcie de pacemaker fiziologic, fiind
denumite din acest motiv i celule P. Acesste celule prezint dimensiuni mai reduse dect
celulele contractile (aproximativ 10 m), se evideniaz optim cu Hematoxilin feric, sunt
alungite, cu o citoplasm mai clar, cu nucleul central, cu miofibrile puine situate la periferie
i organitele slab dezvoltate: rare mitocondrii, RE redus, cteva vezicole pinocitozice
subsarcolemale, fr tubi T, fr elemente ale RS, fr jonciuni de tip strie scalariform, doar
9
HISTOLOGIE GENERAL CORNELIA AMLINEI
desmozomi. Celulele P iniiaz impulsurile prin modificarea spontan a permeabilitii
membranare pentru electrolii, prin deschiderea pompelor ionice, n special a celor de Sodiu.
Aceast depolarizare este meninut activ prin ATP-azele membranare. Ele imprim cel mai
rapid ritm al impulsurilor de contracie: 70 bti/minut. Impulsurile sunt rspndite de-a
lungul celulelor musculare cardiace atriale, determinnd contracia acestora.
Celulele situate n Nodul Atrio-Ventricular (Ashoff-Tawara) prezint celule similare-celule P
care prezint autonomie funcional, putnd iniia impulsuri la o rat de aproximatv jumtate din
ritmul celor din Nodul Sino-Atrial: 35 bti/minut.
Fascicolul atrio-ventricular al lui His, cu ramurile sale i reeaua Purkinje sunt alctuite din
celele Purkinje. Acestea prezint un diametru mai mare dect celulele cardiace comune, sut
globuloase, neregulate ca form, prezint jonciuni intercelulare de tip desmozomi i rare gap.
Sarcoplasma conine o cantitate mare de glicogen, poziionat mai ales central, ceea ce determin
paloarea citoplasmei n coloraiile uzuale. La periferie se gsesc miofibrile. Nucleii (1-2/celul)
sunt rotunjii i mult mai mari dect ai celulelor adiacente. ME relev organite moderat
dezvoltate, lipsa tubilor T, RE slab dezvoltat, mitocondrii numeroase.
ntre celulele Purkinje i celulele contractile apar celulele T (tranziionale) care sunt mai
alungite, ncep s prezinte mai multe organite, mai puin glicogen, fr tubi T, cu miofibrile
ataate la nivelul jonciunilor, cu o orientare aproximativ longitudinal, uor spiralat, ns
numrul lor redus nu reuete s confere un aspect striat. Ele se joncioneaz apoi cu celulele
cardiace contractile, avnd rolul de a transmite impulsul pornit din celulele P i de a opri
impulsurile ectopice premature.
Inervaia celulelor cardiace ale sistemului de conducere este vegetativ, cu rolul de a regla
ritmul contraciilor, nu de a le iniia.
Fibrele parasimpatice (din nervul vag) se termin mai ales n jurul Nodului Sino-Atrial i
Nodului Atrio-Ventricular.
Fibrele simpatice (din ganglionii cervicali superiori) inerveaz Nodul Sino-Atrial i Nodul
Atrio-Ventricular. Fibrele adrenergice realizeaz sinapse en passant, lsnd spaii foarte reduse, de
aproximativ 20 nm ntre cele dou componenete.
Stimulii simpatici determin creterea ratei contraciilor cardiace, parasimpaticul fiind
antagonist cu simpaticul.
Celulele mioendocrine
Celulele mioendocrine au fost descrise n 1968 n ME, de ctre Palade i Jamieson. n
urechiua dreapt atrial s-a observat prezena n celulele cardiace, cu localizare perinuclear, a
unor granule electrono-opace, delimitate de membrane, cu dimensiuni de 0,2-0,4 m. Aceste
granule se asociaz cu abundena complexului Golgi i a RER, fiind mai numeroase n atriul drept,
unde pot ajunge la 600/celul dar putnd fi ntlnite i n atriul stng, n ventricoli i n alte
localizri din organism.
Granulele conin un precursor hormonal cu greutate molecular mare: hormonul
natriuretic atrial, auriculina, atriopeptina care secret prin exocitoz dou polipeptide cu efecte
hormonale:
cardiodilatina sau factorul natriuretic atrial (ANF)
cardionatriuretina sau factorul natriuretic cerebral (brain natriuretic factor-BNF) identic
cu un hormon izolat prima dat n creier.
Efectele acestor hormoni sunt:
Aciune asupra tubilor contori renali favoriznd eliminarea de ap, Sodiu, Potasiu
Aciune antagonist asupra complexului Renin-Aldosteron
Relaxarea musculaturii netede vasculare.
Consecinele aciunii acestor hormoni sunt:
Hipotensiune
Scderea volemiei.
10
HISTOLOGIE GENERAL CORNELIA AMLINEI
n Insuficiena cardiac congestiv, n mod adaptativ, BNF circulant crete, constatnduse o cretere a numrului de granule secretorii n celulele cardiace ventriculare. Stimulul de secreie
este reprezentat de gradul de ntindere a fibrelor musculare atriale, ct i nivelul natremiei i
potasemiei.
esutul muscular neted
esutul muscular neted este involuntar, realizeaz o contracie lent, susinut, ritmic,
fiind sub controlul sistemului nervos vegetativ i al substanelor biologic active.
Histoarhitectonie i organizare
esutul muscular neted prezint modaliti diverse de organizare dup localizarea sa,
putndu-se observa urmtoarele situaii:
celule musculare netede izolate dispuse n esutul conjunctiv al capsulei, septurilor i stromei
prostatei, n capsula i septurile splenice, n esutul subcutanat perineal i penian, n esutul
conjunctiv mamelonar, de-a lungul ductelor lactifere, n intima arterelor i n adventicea venelor
celule izolate, dispuse circumferenial, dispersate de-a lungul arteriolelor precapilare i a
metarteriolelor
celule musculare netede grupate n fascicole fine n muchiul erector al firului de pr asociat
foliculului pilos i glandei sebacee, n muchii intrinseci ai globului ocular, n muchiul
constrictor pupilar al irisului (dispoziie n band inelar), n muchiul dilatator pupilar
(dispoziie radiar), n muchii ciliari, n muchiul Brcke al vilozitii intestinale
celule musculare netede grupate sub forma unei pturi subiri n pielea scrotului (muchiul
dartos)
celule musculare netede grupate ntr-o ptur sau tunic alctuit din celule dispuse ordonat,
circumferenial, n asociere cu esutul conjunctiv n pereii vaselor sanguine (tunica medie), n
pereii organelor tubulare (tractul respirator, ductele mari ale glandelor anexe ale tubului
digestiv-ducte biliare, pancreatice, tractul urinar-calice, bazinet, tractul genital masculin-conuri
eferente, epididim)
celule musculare netede grupate n dou-trei tunici cu orientare diferit (circular sau
longitudinal) n tractul digestiv-din treimea mijlocie a esofagului pn la sfincterul anal intern,
att n musculara mucoasei, ct i n musculara extern (propriu-zis), n trompele uterine, n
ductul deferent, n vezicolele seminale, n ureter
celule musculare netede dispuse sub form de reea de mnunchiuri groase intersectate, asociate
cu un bogat esut conjunctiv-aranjare plexiform, n miometrul uterului i n musculara vezicii
urinare.
Structur i ultrastructur
Celulele musculare netede reprezint subuniti organizatorice denumite tradiional fibre.
Ele au un aspect fuziform, cu o lungime de 20 m, n pereii vaselor, putnd ajunge pn la 200 m
n peretele intestinal, n incidenele secionale longitudinale. Fibrele sunt aranjate paralel astfel nct
extremitatea terminal efilat a unei celule vine n raport cu poriunea dilatat a celulei vecine. n
inciden transversal apar rotund-ovalare sau poligonale, de mrimi variabile.
Sarcolema este nconjurat de o membran bazal bogat n hidrocarbonate i de cantiti
reduse de esut conjunctiv coninnd Fibroblaste. n impregnaia argentic se poate evidenia o reea
delicat reticular strns asociat fiecrei fibre musculare netede.
Sarcoplasma este neted, acidofil n HE, cu depozite PAS + de glicogen.
Nucleii sunt unici, centrali, elongai, n inciden longitudinal. Dac se unesc prin linii
imaginare nucleii aparinnd la dou straturi adiacente de celule musculare netede se obine o linie
n zig-zag. Uneori nucleii prezint aspect pliat (de tirbuon) datorit contraciei, constituind i un
criteriu de difereniere fa de Fibroblaste. Nucleul prezint cromatina nuclear dispus sub forma
unor grmezi reduse de-a lungul marginii interne a membranei nucleare i 1-2 nucleoli. n
11
HISTOLOGIE GENERAL CORNELIA AMLINEI
incidenele transversale, nucleii rotunjii se pot observa doar n seciunile mai mari care trec prin
centrele celulelor.
ME de transmisie evideniaz organitele concentrate perinuclear: cisterne RER, ribozomi,
poliribozomi, REN, Complex Golgi. Restul citoplasmei este ocupat de filamente dispersate de
Actin, de 4-8 nm diametru, asociate cu filamente intermediare, de 8-10 nm diametru i filamente
de Miozin, cu 12-16 nm diametru, fr o aranjare ordonat (reea de tip cristalin). Raportul
sarcoplasm/elemente contractile este n favoarea elementelor contractile.
Filamentele subiri sunt relativ lungi (4,5 m) i conin Actin F format din Actin G,
Tropomiozin i Calmodulin, n loc de Troponin.
Miozina este n stare depolimerizat, fr organizarea microfilamentelor, sub form de
filamente scurte (1,5-2,2 m) izolate, capetele globuloase gsindu-se pe toat ntinderea fibrelor;
prezint trei enzime ATP-azice, cu cte dou izoforme.
Raportul Actin: Miozin este de 12:1 pn la 14:1 fa de 2:1 sau 4:1, n celulele musculare
striate.
Citoscheletul este constituit din structuri lineare sau ramificate n ME de transmisie,
denumite plci de aderen. Acestea sunt electrono-opace, cu lungime de 100-300 nm, cu dou
poziii n interiorul celulei:
submembranare
n masa citoplasmatic.
Din punct de vedere molecular, ele sunt costituite din:
-actinin
vinculin
filamin.
Plcile de aderen reprezint zone de ancorare pentru filamentele subiri i intermediare
(echivalente ale liniilor Z din celulele musculare striate). La nivel subsarcoplasmic realizeaz
contacte n focar care stabilizeaz legturile extracelulare cu laminina i fibronectina, fiind reactive
cu anticorpii pentru vinculin i talin. Plcile de aderen sunt solidarizate printr-o reea
anastomozat de la sarcolem spre interiorul sarcoplasmei, alctuit din urmtoarele filamente
intermediare:
Desmin sau Scheletin (la nivelul viscerelor), reprezentnd proteina major n toate
filamentele intermediare ale tuturor celulelor musculare netede
Distrofina
Vimentina (la nivelul vaselor sanguine).
Sarcoplasma mai conine rari microtubuli.
Nu exist tubi T, ci nfundri ale membranei denumite caveole, asociate cu toate ariile
membranare dintre plcile de aderen. Acestea sunt asemntoare vezicolelor pinocitozice din alte
tipuri de celule. Caveolele conin molecule-receptori pentru Ca++, Na+, K+, Mg++. Caveolele
suplinesc RS n asigurarea Ca++ necesar contraciei, deoarece RS este rudimentar, prezentnd doar
cteva canalicule, din care unul perinuclear, cu mici cantiti de calsechestrin.
Sarcoplasma przint receptori pentru: catecolamine, amine fiziologice i hormoni.
Jonciunile intercelulare sunt de tip gap sau nexus. La nivelul zonelor unde lipsete
membrana bazal se permite conexonilor s realizeze legturi intercelulare.
Inervaia
Inervaia este realizat de neuronii viscerali efereni i afereni ai sistemului nervos
autonom. Fluxul eferent este compus din doi neuroni care fac sinaps ntr-un ganglion simpatic
paravertebral i parasimpatic visceral, astfel nct esutul muscular neted este inervat de neuroni
postganglionari. Sinapsele sunt de tip bouton en passant realiznd un raport de intim vecintate
ntre terminaia vegetativ i memebrana plasmatic a fibrei musculare netede, trecnd de-a lungul
su i realiznd mai multe contacte sinaptice. Butonii conin vezicole sinaptice cu transmitori
neuromusculari:
12
HISTOLOGIE GENERAL CORNELIA AMLINEI
Noradrenalina, Adrenalina care determin contracie prin aciunea pe receptorii 1- 2 i
relaxare prin aciunea pe receptorii , stocate sub forma unor vezicole sinaptice cu un material
granular dens n ME de transmisie
Acid -aminobutiric (GABA), Dopamin stocate n vezicole mari, cu un coninut opac n ME
de transmisie
Acetilcolina, care determin relaxarea muchiului neted, stocat n vezicole sinaptice aparent
goale n ME de transmisie
Amine biologic active: Serotonina, Histamina, Acidul glutamic, Glicina
Peptide: Substana P, Releasing hormones (hormoni de eliberare), Enkefaline
Hormoni secretai de celulele neuro-endocrine ale sistemului neuro-endocrin difuz (DNES):
VIP, Colecistokinina, Neurotensina, Motilina.
Contactul neuro-muscular nu este comparabil cu placa neuromotorie a muchiului striat din mai
multe motive:
Distana considerabil ntre cele dou componente (200 m comparativ cu 10-20 m)
Neuromediatorul eliberat de terminaia nervoas trebuie s difuzeze de-a lungul acestei distane
pentru a ajunge la muchi, astfel nct nu toate celulele musculare sunt expuse direct la
transmitor
n mod similar muchiului cardiac, contracia se transmite de la celul la celul prin jonciuni
gap, astfel nct se obine o o activare coordonat ntr-un mnunchi sau strat muscular.
Mecanismul contraciei
Fibra muscular neted se poate scurta la din lungimea sa, genernd o for pe seciune
transversal comparabil cu cea a muchiului striat dar cu un consum mai redus energetic. Scurtarea
se realizeaz prin mecanismul glisrii filamentelor de Actin, cuplate la cele de Miozin. Scurtarea
considerabli poate fi atribuit lungimii mult mai mari a filamentelor de Actin (4,6 m) comparativ
cu cele din fibra muscular striat (1,6 m).
Ca++ difuzeaz n celul odat cu depolarizarea membranei, fiind eliberat din RS
subplasmalemal i se cupleaz la o protein: Calmodulina. Se realizeaz astfel complexul Ca++Calmodulin n loc de Ca++-Troponin. Complexul Ca++-Calmodulin activeaz ATP-aza
(miozinkinaza) de la capetele moleculei de Miozin, determinnd fosforilarea lanului su uor.
Fosforilarea are loc lent, contracia maxim realizndu-se dup circa un minut.
Clivajul ATP induce o modificare conformaional n capetele Miozinei care determin o
translocaie a filamentelor de Actin adiacente.
Miozina hidrolizeaz ATP-ul la aproximativ 10% din rata existent n celulele musculare
striate scheletale, astfel nct contracia este lent.
Gradul contraciei este influenat de hormoni care acioneaz via AMPc, creterea sa
determinnd activarea lanurilor uoare ale Miozinei, cu fosforilarea acesteia i contracie iar
scderea sa determinnd relaxare. Hormonii sexuali feminini acioneaz prin intermediul acestui
mecanism:
Estrogenii determin creterea nivelului AMPc
Progesteronul are un efect opus.
Exist diferene n activarea muchiului neted din variate esuturi sau organe:
Muchii netezi viscerali funcioneaz n manier sinciial-muchi neted unitar; aceasta se
datoreaz unei inervaii unitare: o fibr vegetativ la mai multe fibre musculare netede; prezint
autoritmicitate; stimulii generai intrinsec sunt condui prin jonciuni gap de la o celul la alta,
ntr-o arie larg muscular care se contract la unison; astfel se produc undele peristaltice
intestinale i undele spontane similare din ureter i ductele biliare; capacitatea de contracie este
atribuit cuplajului electric al fibrelor adiacente, n mod diferit fa de muchiul scheletal unde
fibrele sunt izolate electric, fiecare necesitnd stimularea prin propria sa jonciune neuromuscular.
13
HISTOLOGIE GENERAL CORNELIA AMLINEI
Muchiul neted multiunitar, din pereii arterelor mari, corpii ciliari, iris, pereii ductului
deferent produc contracii foarte precise i gradate; n acetia exist foarte puine evidene ale
conducerii impulsurilor de la o fibr la alta, prin jonciuni gap, fiecare fibr fiind inervat;
contracia este relativ rapid; sinapsele en passant menin n pereii vasculari un status de
contracie parial denumit tonus muscular, care asigur reglarea presiunii hidrostatice, fiind
supus modulrii prin influene hormonale; celulele musculare netede de acest tip prezint
receptori pentru:
Noradrenalin, Angiotensin, Vasopresin (vasoconstrictori)
Bradikinine, Prostaglandine (vasodilatatori).
Factori hormonali implicai n reglarea contraciei
Modificarea nivelului hormonilor Estrogeni din timpul ciclului menstrual se reflect n
variabilitatea coninutului de Actin i Miozin din muchiul neted uterin, Estrogenii determinnd
contracia i Progesteronul determinnd relaxarea.
Hormonii eliberai de Hipofiza posterioar sunt octapeptide care difer n coninutul a doi
aminoacizi, stocai n granule neuro-secretorii ce conin i ATP i o neurofizin-protein de cuplare
legat noncovalent de hormon. ADH/ Vasopresina, n doze mari, determin contracia muchiului
neted din arterele mici i arteriole, iar n doze fiziologice crete permeabilitatea poriunilor distale
ale nefronilor.
Oxitocina determin contracia musculaturii netede uterine, putnd fi utilizat ca inductor
exogen al travaliului. Deasemenea induce contracia celulelor mioepiteliale ale alveolelor secretorii
i ductelor glandei mamare. Secreia oxitocinei este declanat de stimuli nervoi care ajung la
Hipotalamus, aici iniiindu-se un reflex neuro-umoral. La nivelul uterului, stimulul este declanat de
distensia vaginului sau a colului iar la nivelul glandei mamare, de supt.
Heterogenitatea fibrelor musculare netede
Se pot diferenia dou tipuri de fenotipuri fibrelor musculare netede:
Fenotipul contractil, cu o capacitate foarte redus de diviziune
Fenotipul secretor care apare n cursul histogenezei, ca prim fenotip. Acest fenotip se prezint
sub forma celulelor rmuroase, localizate n media pereilor vasculari, a musculaturii
uterine. Ele prezint mai puine filamente contractile, o mai mare dezvoltare a organitelor
implicate n sintez (RER, complex Golgi perinucleare), jonciuni intercelulare de tip miomiocitar, prin care i coordoneaz procesele de secreie. Celulele rmuroase sunt principalele
celule responsabile pentru sinteza matricei esutului conjunctiv: Proteoglicani, Colagen tip III,
II, Fibronectin, Laminin i a Elastinei i Colagenului tip IV (n membranele bazale). Ele i
menin capacitatea de proliferare.
n esutul subendotelial al Intimei arteriale se gsesc celule miointimale care prezint
ambele fenotipuri, contractil i secretor, cu urmtoarele proprieti:
prezint capaciti fagocitare, putnd deveni, prin fagocitoza lipidelor (Colesterolului), ca i
Macrofagele, celule spumoase
produc elemente de matrice, factori de cretere, factori chemotactici, prostacicline.
Tipuri particulare de fibre musculare netede
Celulele juxtaglomerulare sunt celule musculare din media arteriolelor aferente sau uneori
eferente glomerulare care vin n contact cu tubul contort distal, n dreptul Maculei Densa. Sunt
celule musculare netede modificate, mai globuloase, cu nuclei sferici centrali, cu caracter epitelioid,
cu granule de secreie PAS+, delimitate de membrane, cu coninut electronodens. n aceast zon
limitanta elastic intern dispare, ca i esutul subendotelial, astfel nct celulele juxtaglomerulare
sunt aproape de lumen, funcionnd ca baroreceptori.
Ele secret Renin, Eritropoietin, Kalicrein, Prostaglandine. Renina transform
Angiotensinogenul prin hidroliz n Angiotensin I (decapeptid) care este transformat de o enzim
de conversie (secretat de endoteliul pulmonar) n Angiotensin II (octapeptid activ) care este
puternic vasoconstrictoare.
14
HISTOLOGIE GENERAL CORNELIA AMLINEI
Inhibitorii enzimei de conversie de tipul Captoprilului, Enalaprilului sunt utilizai pentru
tratarea Bolii hipertensive cronice.
Renina stimuleaz zona glomerulat a Corticosuprarenalei s sintetizeze i s elibereze
Aldosteronul care stimuleaz resorbia de Na+ i ap la nivelul tubilor contori distali, crescnd
astfel volumul i presiunea sanguin. Aldosteronul este i un potent vasoconstrictor.
Stimulii eliberrii de Renin sunt: scderea volumului sanguin, scderea concentraiei Na+.
Celulele Maculei densa monitorizeaz concentraia NaCl n arteriola aferent i regleaz
eliberarea de Renin din celulele juxtaglomerulare ntr-o manier paracrin.
Celulele mioide testiculare sunt situate n jurul tubilor seminiferi. Prezint o ultrastructur
asemntoare fibrelor musculare netede, citoscheletul coninnd filamente de Desmin. Imprim
micri peristaltice tubilor seminiferi n vederea transportului spermatozoizilor.
Capacitatea de noire, reparare i difereniere
Celulele musculare netede sunt capabile de diviziune pentru a se menine sau pentru a crete
numeric, n special cele cu fenotip secretor. Ele rspund la distrugeri prin mitoze.
Celulele musculare netede ale miometrului prolifereaz n timpul ciclului menstrual i al
sarcinii, sub control hormonal.
Fibrele musculare netede vasculare se divid n mod regulat la adult pentru a nlocui celulele
distruse sau mbtrnite. Fibrele musculare netede pot lua natere prin diviziune i diferenierea
celulelor endoteliale i a pericitelor, mai ales la vasele n curs de dezvoltare.
Procesele de Hipertrofie-Hiperplazie-Metaplazie
Hipertrofia de efort reprezint o mrire a fiecrei fibre musculare netede datorit apariiei
unor obstacole de-a lungul unui organ tubular. n amonte de obstacol se va produce hipertrofia
fibrelor musculare netede din perete.
Fibrele musculare netede ale miometrului au o lungime de 40-80 m, n uterul negestant i
pot ajunge la 500-600 m n uterul gestant. Acest fenomen se datoreaz suprapunerii a trei
mecanisme:
Hipertrofie propriu-zis: celulele musculare netede cresc de aproximativ 10 ori ca dimensiuni
Hiperplazie, prin multiplicarea numrului de celule musculare prin mitoze n special ale unor
miocite de tip secretor care se vor transforma ulterior n miocite cu fenotip contractil
Metaplazie prin diferenierea Miofibroblastelor spre miocite contractile.
Aceste modificri sunt hormono-dependente. Dup gestaie, datorit prbuirii nivelelor
hormonale, se stimuleaz genele apoptotice a celulelor musculare netede i a Miofibroblastelor cu
revenirea spre dimensiunile iniiale.
Progesteronul i Relaxina mpiedic contracia i Estrogenii materni i corticosteroizii fetali
stimuleaz contraciile miometrului.
Aspecte patologice
Leiomiofibromul reprezint proliferarea miometrului sub stimulul estrogenic.
Musculatura neted vascular poate suferi procese de metaplazie condroid sau
fibroblastic.
15
HISTOLOGIE GENERAL CORNELIA AMLINEI
BIBLIOGRAFIE
1. Amlinei C.; Histologie general, Ed. Corson, Iai, Romnia, 2001.
2. Amlinei C., Balan R., Cotuiu C.; Histology-microscopic diagnosis of organs, Ed. Cantes, Iai,
Romnia, 2000.
3. Bdescu A., Cruntu I., Amlinei C., Floarea-Strat A., Adomnici M.; esuturi normale-curs de
Histologie, Ed. Graphix, Iai, Romnia, 1994.
4. Burkitt H. G., Young B., Heath J. W., Deakin P. J.; Wheaters Functional Histology-a text and atlas, Ed.
Churchilll Livingstone, New York, U.S.A., 1995.
5. Cruntu I. D., Cotuiu C.; Histologie special-ghid pentru lucrri practice, Ed. Apollonia, Iai, 1998.
6. Fawcett D. W., Bloom W.; A textbook of Histology, Ed. Chapman & Hall, New York, U.S.A., 1994.
7. Gartner P. L., Hiatt L. J., Strum M. J.; Cell Biology and Histology, Ed. Williams & Wilkins, Baltimore,
Maryland, 1998.
8. Junqueira L. C., Carneiro J., Kelley O. R.; Basic Histology, Ed. Appleton & Lange, Stamford,
Connecticut, U.S.A., 1995.
9. Poirier J., Ribadeau-Dumas J. L., Catala M., Gherardi R. K., Bernaudin J. F.; Histologie molculaire,
Ed. Masson, Paris, France, 1997.
10. Ross H. M., Romrell J. L., Kaye J. G.; Histology-a text and atlas, Ed. Williams & Wilkins, Baltimore,
Maryland, 1995.
16
S-ar putea să vă placă și
- Tesut Muscular Amalinei PDFDocument16 paginiTesut Muscular Amalinei PDFbDashÎncă nu există evaluări
- Tesut Muscular 01Document10 paginiTesut Muscular 01Alina Alexandra ToeaÎncă nu există evaluări
- Tesutul MuscularDocument4 paginiTesutul MuscularAriela Săvuţ100% (1)
- Curs Histo 7 Tesuturi MusculareDocument12 paginiCurs Histo 7 Tesuturi MusculareAndrada ManguÎncă nu există evaluări
- Fibra MuscularaDocument39 paginiFibra MuscularaIuliana Alexandra Valcu100% (3)
- Tesutul MuscularDocument3 paginiTesutul Muscularkty_Încă nu există evaluări
- Tesuturi MusculateDocument40 paginiTesuturi MusculateIoana NavaliciÎncă nu există evaluări
- Curs 6 ŢESUTURILE MUSCULARE !Document20 paginiCurs 6 ŢESUTURILE MUSCULARE !Anonymous foLBZlRAeK100% (1)
- Tesut Muscular Curs 2019 2020Document10 paginiTesut Muscular Curs 2019 2020rares gherghinaÎncă nu există evaluări
- Fiziologie LP NR 3 Anul IDocument13 paginiFiziologie LP NR 3 Anul IAnna MariaÎncă nu există evaluări
- Structura Tesutului Muscular 3.1Document8 paginiStructura Tesutului Muscular 3.1ligiailonaÎncă nu există evaluări
- Tesut Muscular CursDocument6 paginiTesut Muscular CursVictor Aurel StefanescuÎncă nu există evaluări
- Ţesuturile MusculareDocument27 paginiŢesuturile MusculareAndreea PanaitiuÎncă nu există evaluări
- Cursul IV, Note de Curs Neuro-MuscularDocument16 paginiCursul IV, Note de Curs Neuro-MuscularYusef Ceva100% (1)
- Referat Tesut MuscularDocument10 paginiReferat Tesut Muscularmel1gÎncă nu există evaluări
- Tesuturi MusculareDocument19 paginiTesuturi MusculareCarmen Ovidiu MihaiÎncă nu există evaluări
- Structura Tesutului MuscularDocument8 paginiStructura Tesutului MuscularVass GeosanuÎncă nu există evaluări
- Tesutul MuscularDocument8 paginiTesutul MusculardanÎncă nu există evaluări
- Structura Tesutului MuscularDocument8 paginiStructura Tesutului MuscularAna-Maria TanasăÎncă nu există evaluări
- 7 Tesuturi MusculareDocument9 pagini7 Tesuturi MusculareValentin MoisăÎncă nu există evaluări
- Curs 4 - CitoscheletDocument46 paginiCurs 4 - CitoscheletAdrian UrsuÎncă nu există evaluări
- Fibra Musculară Striată (FMS)Document27 paginiFibra Musculară Striată (FMS)Teodora PeicaÎncă nu există evaluări
- Tesut Muscular 26324Document111 paginiTesut Muscular 26324Galina BotnariÎncă nu există evaluări
- Organul EfectorDocument9 paginiOrganul EfectorMary.GeorgianaÎncă nu există evaluări
- FiziologieDocument47 paginiFiziologieGeorgian SerafimÎncă nu există evaluări
- 5 - MuscularDocument14 pagini5 - MuscularVioleta GeorgescuÎncă nu există evaluări
- Curs - Muschi StriatDocument40 paginiCurs - Muschi StriatDiana Didi Dia100% (1)
- Curs 8 Tesutul Muscular PPTDocument29 paginiCurs 8 Tesutul Muscular PPTValiDrancaÎncă nu există evaluări
- Muscular MD RO 2018-2019Document49 paginiMuscular MD RO 2018-2019Crystyan Crysty CojocaruÎncă nu există evaluări
- Curs 9 Tesuturi MusculareDocument11 paginiCurs 9 Tesuturi MusculareAndra GarbeaÎncă nu există evaluări
- HistologieDocument8 paginiHistologieLorena DamianÎncă nu există evaluări
- Curs Nr. 5 Cit - NouDocument53 paginiCurs Nr. 5 Cit - NouLaura HutzelÎncă nu există evaluări
- Curs II - Muschiul NetedDocument8 paginiCurs II - Muschiul NetedSimona Balasescu100% (2)
- Curs 8 HistologieDocument12 paginiCurs 8 HistologieMinigirl AlexandraÎncă nu există evaluări
- Muschi NetedDocument6 paginiMuschi Netediuuu3Încă nu există evaluări
- Muşchiul Şi Contracţia MuscularăDocument9 paginiMuşchiul Şi Contracţia MuscularăIonut CojoÎncă nu există evaluări
- Forma Si Structura MuschilorDocument3 paginiForma Si Structura MuschilorTepeş-Stroia EugenÎncă nu există evaluări
- MuschiDocument49 paginiMuschiSANDY ATTIA GURGIS BISHAYÎncă nu există evaluări
- Biologie Celulara Si HistologieDocument78 paginiBiologie Celulara Si HistologieRaluca StefanescuÎncă nu există evaluări
- Fiziologia MuşchilorDocument74 paginiFiziologia MuşchilorXiusa TurcanÎncă nu există evaluări
- Sistemul MuscularDocument30 paginiSistemul Muscularmidnightpoison1Încă nu există evaluări
- Bio ChimDocument22 paginiBio ChimEu VictorÎncă nu există evaluări
- Histologie - Tesutul MuscularDocument30 paginiHistologie - Tesutul MuscularIoan AsztalosÎncă nu există evaluări
- Sistemul NeuromuscDocument67 paginiSistemul NeuromuscAlexandru PurcelÎncă nu există evaluări
- Curs 2 InimaDocument20 paginiCurs 2 InimaAttila SzakacsÎncă nu există evaluări
- Biofizica Aparatului LocomotorDocument10 paginiBiofizica Aparatului LocomotorAurel BoleaÎncă nu există evaluări
- Referat Anatomie 3Document6 paginiReferat Anatomie 3Mihnea BucurÎncă nu există evaluări
- Lp-4-Forme CelulareDocument7 paginiLp-4-Forme CelulareLucia RaluÎncă nu există evaluări
- Ţesut MuscularDocument12 paginiŢesut MuscularStan DianaÎncă nu există evaluări
- Tesutul MuscularDocument8 paginiTesutul Muscularbianca100% (1)
- Curs 4 FiziologieDocument10 paginiCurs 4 FiziologieDAROIU REBECAGEORGIANAÎncă nu există evaluări
- Structura Morfologica A CarniiDocument7 paginiStructura Morfologica A CarniiCapatina Victor OctavianÎncă nu există evaluări
- MuscularDocument46 paginiMuscularCatalin CusnirÎncă nu există evaluări
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 4 din 5 stele4/5 (8)
- Manual Utilizare RodiabDocument17 paginiManual Utilizare RodiabTamaraBmBÎncă nu există evaluări
- Elena Avramescu - Bazele Anatomice Ale MiscariiDocument108 paginiElena Avramescu - Bazele Anatomice Ale Miscariiiri0100% (10)
- Vascularizatie MB SuperiorDocument3 paginiVascularizatie MB SuperiorNic NicÎncă nu există evaluări
- Actiune Muschii Membrului SuperiorDocument21 paginiActiune Muschii Membrului SuperiorTamaraBmBÎncă nu există evaluări
- Nutritia InimiiDocument9 paginiNutritia InimiiAna Maria CiurdoreanÎncă nu există evaluări