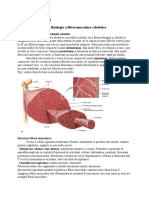
S-ar putea să vă placă și
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 4 din 5 stele4/5 (8)
- Microcirculația sanguină: Următoarea generație de asistență socialăDe la EverandMicrocirculația sanguină: Următoarea generație de asistență socialăÎncă nu există evaluări
- Padurile EcuatorialeDocument4 paginiPadurile EcuatorialeElena Botezatu92% (12)
- Mecanismul Contractiilor MusculareDocument18 paginiMecanismul Contractiilor MusculareMoisiiMihail100% (2)
- Secusa. Tetanosul Oboseala Musculara - CompletDocument22 paginiSecusa. Tetanosul Oboseala Musculara - CompletEnache Aniela100% (1)
- Tesut Muscular Curs 2019 2020Document10 paginiTesut Muscular Curs 2019 2020rares gherghinaÎncă nu există evaluări
- Pădurile TropicaleDocument7 paginiPădurile TropicaleAdrian TodericaÎncă nu există evaluări
- FiziologieDocument47 paginiFiziologieGeorgian SerafimÎncă nu există evaluări
- Fibra MuscularaDocument39 paginiFibra MuscularaIuliana Alexandra Valcu100% (3)
- Marile Descoperiri GeograficeDocument31 paginiMarile Descoperiri GeograficeXiusa Turcan100% (1)
- MuscularDocument46 paginiMuscularCatalin CusnirÎncă nu există evaluări
- 06 Contractia Musculara 2017-12-04Document11 pagini06 Contractia Musculara 2017-12-04costica16100% (1)
- Biofizica Contractiei MusculareDocument6 paginiBiofizica Contractiei MusculareAndrone CameliaÎncă nu există evaluări
- Romanii in Lupta AntiotomanaDocument45 paginiRomanii in Lupta AntiotomanaXiusa Turcan100% (1)
- Sarcomerul CardiacDocument17 paginiSarcomerul CardiacCiobotaru AlexandraÎncă nu există evaluări
- Ideologii În Epoca ModernăDocument10 paginiIdeologii În Epoca ModernăXiusa Turcan100% (1)
- Muşchiul Şi Contracţia MuscularăDocument9 paginiMuşchiul Şi Contracţia MuscularăIonut CojoÎncă nu există evaluări
- Fiziologia Contractiei MusculareDocument4 paginiFiziologia Contractiei MusculareIoana irimiaÎncă nu există evaluări
- Referat FiziollogieDocument7 paginiReferat FiziollogiedarlaloredanaÎncă nu există evaluări
- Structura Tesutului MuscularDocument8 paginiStructura Tesutului MuscularVass GeosanuÎncă nu există evaluări
- Biofizica Contracţiei MusculareDocument6 paginiBiofizica Contracţiei MuscularenoruletÎncă nu există evaluări
- Morfologia Muschilor Striati 1Document10 paginiMorfologia Muschilor Striati 1AlexColibaÎncă nu există evaluări
- Contractia MuscularaDocument4 paginiContractia MuscularaTudor OlteanÎncă nu există evaluări
- Tesutul MuscularDocument4 paginiTesutul MuscularAriela Săvuţ100% (1)
- Tesut Muscular 26324Document111 paginiTesut Muscular 26324Galina BotnariÎncă nu există evaluări
- Sistemului NeuromuscularDocument54 paginiSistemului NeuromuscularStefi MaryÎncă nu există evaluări
- Biofizica Contractiei MusculareDocument11 paginiBiofizica Contractiei MusculareMircea CiobîcăÎncă nu există evaluări
- Bio ChimDocument22 paginiBio ChimEu VictorÎncă nu există evaluări
- FIziologia Aparatului NeuromuscularDocument14 paginiFIziologia Aparatului NeuromuscularLoredana Boghez100% (1)
- Tesut Muscular Amalinei PDFDocument16 paginiTesut Muscular Amalinei PDFbDashÎncă nu există evaluări
- Curs - Muschi StriatDocument40 paginiCurs - Muschi StriatDiana Didi Dia100% (1)
- Sistemul NeuromuscDocument67 paginiSistemul NeuromuscAlexandru PurcelÎncă nu există evaluări
- Contracția Musculara BF FIMDocument16 paginiContracția Musculara BF FIMVlad ȘtefanÎncă nu există evaluări
- CURS MUSCHI FIZIO PPTDocument46 paginiCURS MUSCHI FIZIO PPTIrina MureșanÎncă nu există evaluări
- Structura Tesutului MuscularDocument8 paginiStructura Tesutului MuscularAna-Maria TanasăÎncă nu există evaluări
- Curs 5 Muschiul Striat NetedDocument6 paginiCurs 5 Muschiul Striat Netedp3tru_mzqÎncă nu există evaluări
- Fiziologie LP NR 3 Anul IDocument13 paginiFiziologie LP NR 3 Anul IAnna MariaÎncă nu există evaluări
- Structura Si Proprietetile Fiziologice Ale Fibrelor Muschilor StriatiDocument2 paginiStructura Si Proprietetile Fiziologice Ale Fibrelor Muschilor StriatiDaniela IstratiÎncă nu există evaluări
- Cursul IV, Note de Curs Neuro-MuscularDocument16 paginiCursul IV, Note de Curs Neuro-MuscularYusef Ceva100% (1)
- Biologie Celulara Si HistologieDocument78 paginiBiologie Celulara Si HistologieRaluca StefanescuÎncă nu există evaluări
- Tesut Muscular 01Document10 paginiTesut Muscular 01Alina Alexandra ToeaÎncă nu există evaluări
- Biofizica Contracţiei MusculareDocument4 paginiBiofizica Contracţiei Muscularechamps2009Încă nu există evaluări
- 7 Tesuturi MusculareDocument9 pagini7 Tesuturi MusculareValentin MoisăÎncă nu există evaluări
- Fiziologie TotDocument34 paginiFiziologie TotbamsweÎncă nu există evaluări
- Curs 5 BiofizicaDocument9 paginiCurs 5 BiofizicacurteanancaÎncă nu există evaluări
- Sistemul MuscularDocument9 paginiSistemul MuscularDiana AndreeaÎncă nu există evaluări
- Curs Fiziologia Mușchilor Striați Și Netezi-34826Document57 paginiCurs Fiziologia Mușchilor Striați Și Netezi-34826Alina IlovanÎncă nu există evaluări
- Biofizica Contractiei Musculare Sistemul Muscular PDFDocument1 paginăBiofizica Contractiei Musculare Sistemul Muscular PDFandamuresan3Încă nu există evaluări
- Biofizica Aparatului LocomotorDocument10 paginiBiofizica Aparatului LocomotorAurel BoleaÎncă nu există evaluări
- Structura Țesutului Muscular, Compoziția Chimică .Document21 paginiStructura Țesutului Muscular, Compoziția Chimică .Iuliu DigorÎncă nu există evaluări
- Citoplasma CelularaDocument16 paginiCitoplasma CelularaEnea DanielaÎncă nu există evaluări
- C4 - NeuroMuscular PDFDocument41 paginiC4 - NeuroMuscular PDFAndrei CiobotaruÎncă nu există evaluări
- Curs 4 - CitoscheletDocument46 paginiCurs 4 - CitoscheletAdrian UrsuÎncă nu există evaluări
- Curs 5Document14 paginiCurs 5sham harbaÎncă nu există evaluări
- Curs 2 InimaDocument20 paginiCurs 2 InimaAttila SzakacsÎncă nu există evaluări
- Tesuturi Excitabile IIIDocument5 paginiTesuturi Excitabile IIINirca VadimÎncă nu există evaluări
- Curs 6, 7, 8Document64 paginiCurs 6, 7, 8Marian Dan IngridÎncă nu există evaluări
- Curs 2Document29 paginiCurs 2Marian Dan IngridÎncă nu există evaluări
- Partea IIIDocument35 paginiPartea IIIAlina TeodosieÎncă nu există evaluări
- Curs 4Document31 paginiCurs 4Marian Dan IngridÎncă nu există evaluări
- Prim AjutorDocument36 paginiPrim AjutorMarian Dan IngridÎncă nu există evaluări
- Prim AjutorDocument49 paginiPrim AjutorMarian Dan Ingrid100% (1)
- DigestivDocument15 paginiDigestivXiusa TurcanÎncă nu există evaluări
- Limba Română Și Artă FotograficăDocument6 paginiLimba Română Și Artă FotograficăXiusa TurcanÎncă nu există evaluări
- Bio TestDocument7 paginiBio TestXiusa TurcanÎncă nu există evaluări
- EnergiaDocument7 paginiEnergiaXiusa TurcanÎncă nu există evaluări
- Bio Test Uman RoDocument6 paginiBio Test Uman RoXiusa TurcanÎncă nu există evaluări
- Constantele Eritrocitului NormalDocument37 paginiConstantele Eritrocitului NormalXiusa TurcanÎncă nu există evaluări
- NeuronDocument70 paginiNeuronXiusa TurcanÎncă nu există evaluări
- Test Biologia UmanistDocument4 paginiTest Biologia UmanistXiusa TurcanÎncă nu există evaluări
- Mark Finley 10 ZileDocument45 paginiMark Finley 10 ZileXiusa Turcan100% (1)
- Numere Intregi in Viata MeaDocument4 paginiNumere Intregi in Viata MeaXiusa Turcan0% (1)
- PestisiamfibieniDocument8 paginiPestisiamfibieniOlga RotariÎncă nu există evaluări
- EnergiaDocument7 paginiEnergiaXiusa TurcanÎncă nu există evaluări
- 1 Decembrie, Ziua Naţională A RomânieiDocument4 pagini1 Decembrie, Ziua Naţională A RomânieiXiusa TurcanÎncă nu există evaluări
- Evaluare La Lupta Antiotoman (Document3 paginiEvaluare La Lupta Antiotoman (Xiusa TurcanÎncă nu există evaluări
- Evaluare Economicul Si Socialul MedievalDocument3 paginiEvaluare Economicul Si Socialul MedievalXiusa TurcanÎncă nu există evaluări
- Crearea Unui SiteDocument1 paginăCrearea Unui SiteXiusa TurcanÎncă nu există evaluări
- ReptileDocument34 paginiReptileOlga RotariÎncă nu există evaluări