S-ar putea să vă placă și
- Biofizica Contracţiei MusculareDocument6 paginiBiofizica Contracţiei MuscularenoruletÎncă nu există evaluări
- 2-Biofizica Contractiei MusculareDocument26 pagini2-Biofizica Contractiei MusculareLuiza Anghel DutaÎncă nu există evaluări
- Proprietatile NervilorDocument2 paginiProprietatile NervilorIonica RusescuÎncă nu există evaluări
- Sindromul Piramidal - de Neuron Motor CentralDocument14 paginiSindromul Piramidal - de Neuron Motor CentralMally FoxÎncă nu există evaluări
- A 7a Biologie Planificarea Caledaristica Si Pe Unitati 2017Document42 paginiA 7a Biologie Planificarea Caledaristica Si Pe Unitati 2017Simona LunguÎncă nu există evaluări
- Subiecte Referate Biofizica 2011Document4 paginiSubiecte Referate Biofizica 2011Onutza IoanaÎncă nu există evaluări
- Prezentare ScoalaDocument21 paginiPrezentare ScoalaIzabela Andrea100% (1)
- Margutoiu Alexandru - Sistemul Iritativ VegetativDocument4 paginiMargutoiu Alexandru - Sistemul Iritativ VegetativMargutoiu AlexandruÎncă nu există evaluări
- 0 Plan 6Document9 pagini0 Plan 6Ciucurel Ana VeronicaÎncă nu există evaluări
- Malformatii CraniofacialeDocument55 paginiMalformatii CraniofacialeCristina CoşulăÎncă nu există evaluări
- Fiziologia NeuronuluiDocument4 paginiFiziologia NeuronuluiRaluca MidoschiÎncă nu există evaluări
- 2 - HumerusDocument13 pagini2 - HumerusJim Boy100% (1)
- Curs 6 ŢESUTURILE MUSCULARE !Document20 paginiCurs 6 ŢESUTURILE MUSCULARE !Anonymous foLBZlRAeK100% (1)
- Celula, Proprietatile CeluleiDocument12 paginiCelula, Proprietatile CeluleiAlexÎncă nu există evaluări
- Controlul HomeostazieiDocument5 paginiControlul HomeostazieiDaiana RotaruÎncă nu există evaluări
- Proiect de Lectie - Cavitatea BucalăDocument5 paginiProiect de Lectie - Cavitatea BucalăAndreea RobuÎncă nu există evaluări
- ANALIZATORIIDocument10 paginiANALIZATORIIBosneaga DanielaÎncă nu există evaluări
- Diencefalul Talamus CursDocument9 paginiDiencefalul Talamus Cursmimi12345678910Încă nu există evaluări
- Curs Proteazomi s2Document80 paginiCurs Proteazomi s2Simona BalasescuÎncă nu există evaluări
- Curriculum La Biologie, Clasele VI-IXDocument21 paginiCurriculum La Biologie, Clasele VI-IXLiusika RotariÎncă nu există evaluări
- Histologie Examen RaspunsuriDocument33 paginiHistologie Examen RaspunsuriMarin HarabagiuÎncă nu există evaluări
- Fractura FEMUR (Roman (Mircea) Elena Mihaela)Document7 paginiFractura FEMUR (Roman (Mircea) Elena Mihaela)Ruxandra RaduÎncă nu există evaluări
- Neuro MotorDocument6 paginiNeuro MotorAnk GeorgyÎncă nu există evaluări
- ERR Teor Inj. IntramuscularăDocument4 paginiERR Teor Inj. IntramuscularăLarisa PalamariÎncă nu există evaluări
- Explorarea Sistemului CirculatorDocument206 paginiExplorarea Sistemului CirculatorPopa D. SilviuÎncă nu există evaluări
- AtentiaDocument3 paginiAtentiaFerenkÎncă nu există evaluări
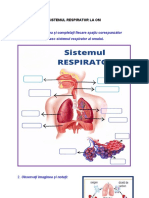
- Observați Imaginea Și Completați Fiecare Spațiu Corespunzător Organelor Ce Alcătuiesc Sistemul Respirator Al OmuluiDocument2 paginiObservați Imaginea Și Completați Fiecare Spațiu Corespunzător Organelor Ce Alcătuiesc Sistemul Respirator Al OmuluiDana MaryÎncă nu există evaluări
- 5 Obiective OperationaleDocument2 pagini5 Obiective OperationaleAndreea PetreaÎncă nu există evaluări
- Referat EnglezaDocument8 paginiReferat EnglezaSerban CristinaÎncă nu există evaluări
- Patologia Pensionarii Si Integrarea Varstnicului in Institutii SpecializateDocument10 paginiPatologia Pensionarii Si Integrarea Varstnicului in Institutii SpecializateioanÎncă nu există evaluări
- Plan de Lectie Limba Budurca ValentinaDocument11 paginiPlan de Lectie Limba Budurca ValentinaValentina BudurcaÎncă nu există evaluări
- Subiecte Referat Amg 1Document2 paginiSubiecte Referat Amg 1Giani AndraÎncă nu există evaluări
- Sindromul KlinefelterDocument7 paginiSindromul Klinefelteranastasieigeorge4962Încă nu există evaluări
- Studiu de Caz Plan de Ingrijire IRADocument13 paginiStudiu de Caz Plan de Ingrijire IRAMIHAELA BRÂNZAC CAS HUŢULEACÎncă nu există evaluări
- Anatomia Si Fiziologia Etajului Inferior Al Tubului DigestivDocument10 paginiAnatomia Si Fiziologia Etajului Inferior Al Tubului DigestivSergiu HelciugÎncă nu există evaluări
- Investigatii Ale Functiei PulmonareDocument16 paginiInvestigatii Ale Functiei PulmonareElena TudoseÎncă nu există evaluări
- Aparatul AuditivDocument4 paginiAparatul AuditivAnne BarbuÎncă nu există evaluări
- Vascozitatea SangeluiDocument21 paginiVascozitatea SangeluiFlorina Grumezescu100% (2)
- 2-Biofizica Contractiei MusculareDocument44 pagini2-Biofizica Contractiei MusculareAlex Pop100% (1)
- Curs 1 Pneumologie-Anatomie OanaDocument59 paginiCurs 1 Pneumologie-Anatomie OanaOana-Gianina NicolaiÎncă nu există evaluări
- Pipp32 - Puericultura - Igiena-Unitatea 3Document8 paginiPipp32 - Puericultura - Igiena-Unitatea 3Ioana MarinÎncă nu există evaluări
- Organite Nedelimitate de EndomembraneDocument37 paginiOrganite Nedelimitate de Endomembranelaur112233Încă nu există evaluări
- Foaie de Observaţie Clinică A BolnavilorDocument23 paginiFoaie de Observaţie Clinică A BolnavilorCarolina Gîrneț - CazacuÎncă nu există evaluări
- Echilibrul OsmoticDocument33 paginiEchilibrul OsmoticOana Rosca100% (1)
- Sistemul MuscularDocument34 paginiSistemul MuscularIleana AndreeaÎncă nu există evaluări
- 10 Fisa de Lucru 3Document4 pagini10 Fisa de Lucru 3apetrei denis gianiÎncă nu există evaluări
- Histologie 1 Curs 2) Epitelii GlandulareDocument6 paginiHistologie 1 Curs 2) Epitelii GlandulareMariaMaryaÎncă nu există evaluări
- Plan de Lectie RECAPITULARE Med InternaDocument4 paginiPlan de Lectie RECAPITULARE Med InternaMARIANA LOREDANA POPESCUÎncă nu există evaluări
- Meningitele Acute Bacteriene-Doctorat IubuDocument34 paginiMeningitele Acute Bacteriene-Doctorat IubuDeli MushroomsÎncă nu există evaluări
- Accident Vascular Cerebral IschemicDocument14 paginiAccident Vascular Cerebral IschemicCarmen SicoeÎncă nu există evaluări
- Biofizica Contractiei MusculareDocument11 paginiBiofizica Contractiei MusculareMircea CiobîcăÎncă nu există evaluări
- Biofizica Contractiei Musculare Sistemul Muscular PDFDocument1 paginăBiofizica Contractiei Musculare Sistemul Muscular PDFandamuresan3Încă nu există evaluări
- Curs 5 BiofizicaDocument9 paginiCurs 5 BiofizicacurteanancaÎncă nu există evaluări
- Biofizica Contracţiei MusculareDocument4 paginiBiofizica Contracţiei Muscularechamps2009Încă nu există evaluări
- 06 Contractia Musculara 2017-12-04Document11 pagini06 Contractia Musculara 2017-12-04costica16100% (1)
- Morfologia Muschilor Striati 1Document10 paginiMorfologia Muschilor Striati 1AlexColibaÎncă nu există evaluări
- Structura Si Proprietetile Fiziologice Ale Fibrelor Muschilor StriatiDocument2 paginiStructura Si Proprietetile Fiziologice Ale Fibrelor Muschilor StriatiDaniela IstratiÎncă nu există evaluări
- Sarcomerul CardiacDocument17 paginiSarcomerul CardiacCiobotaru AlexandraÎncă nu există evaluări
- ElectromiogramaDocument6 paginiElectromiogramaAna Maria100% (1)
- Curs - Muschi StriatDocument40 paginiCurs - Muschi StriatDiana Didi Dia100% (1)
- ColorareaDocument8 paginiColorareaAndrone CameliaÎncă nu există evaluări
- Perceptia VizualaDocument16 paginiPerceptia VizualaAndrone CameliaÎncă nu există evaluări
- Biofizica Receptiei AuditiveDocument14 paginiBiofizica Receptiei AuditiveAndrone Camelia100% (1)
- 05 Curs Receptie VizualaDocument17 pagini05 Curs Receptie VizualaLoveMed09Încă nu există evaluări
- Transport Membranar CURSDocument8 paginiTransport Membranar CURSCathyÎncă nu există evaluări
- Fenomene BioelectriceDocument11 paginiFenomene BioelectriceGeorgeNecoarăÎncă nu există evaluări
- Secretia Pancreatica+bilaDocument42 paginiSecretia Pancreatica+bilaAndrone CameliaÎncă nu există evaluări
- Reglarea Secretiei GastriceDocument23 paginiReglarea Secretiei GastriceAndrone CameliaÎncă nu există evaluări
- Apa in OrganismDocument42 paginiApa in OrganismAndrone Camelia100% (3)
- Curs Reglarea Echilibrului HidricDocument40 paginiCurs Reglarea Echilibrului HidricAndrone CameliaÎncă nu există evaluări
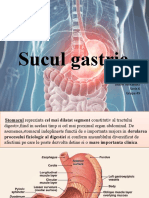
- Sucul GastricDocument46 paginiSucul GastricAndrone CameliaÎncă nu există evaluări
- Compartimente Hidrice FinalDocument31 paginiCompartimente Hidrice FinalAndrone CameliaÎncă nu există evaluări
- Secretia Gastrica FinalDocument42 paginiSecretia Gastrica FinalAndrone CameliaÎncă nu există evaluări
- SalivaDocument70 paginiSalivaAndrone CameliaÎncă nu există evaluări
- Fizio MuscularDocument11 paginiFizio MuscularAndrone CameliaÎncă nu există evaluări
- PoligramaDocument12 paginiPoligramaAndrone CameliaÎncă nu există evaluări
- Sucul GastricDocument46 paginiSucul GastricAndrone CameliaÎncă nu există evaluări
- Ruptura de SplinaDocument19 paginiRuptura de SplinaAndrone CameliaÎncă nu există evaluări
- Fizio MuscularDocument11 paginiFizio MuscularAndrone CameliaÎncă nu există evaluări
- Perceptia VizualaDocument16 paginiPerceptia VizualaAndrone CameliaÎncă nu există evaluări
- Ce Te Face Un Bun Om de EchipaDocument2 paginiCe Te Face Un Bun Om de EchipaAndrone CameliaÎncă nu există evaluări
- Antidepresive ProiectDocument6 paginiAntidepresive ProiectAndrone Camelia100% (1)
- O Scrisoare PierdutaDocument13 paginiO Scrisoare PierdutaAndrone CameliaÎncă nu există evaluări