S-ar putea să vă placă și
- Mecanismul Contractiilor MusculareDocument18 paginiMecanismul Contractiilor MusculareMoisiiMihail100% (2)
- Fibra MuscularaDocument39 paginiFibra MuscularaIuliana Alexandra Valcu100% (3)
- 06 Contractia Musculara 2017-12-04Document11 pagini06 Contractia Musculara 2017-12-04costica16100% (1)
- Biofizica Contractiei MusculareDocument6 paginiBiofizica Contractiei MusculareAndrone CameliaÎncă nu există evaluări
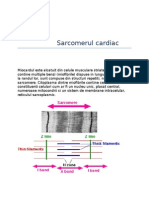
- Sarcomerul CardiacDocument17 paginiSarcomerul CardiacCiobotaru AlexandraÎncă nu există evaluări
- MuscularDocument9 paginiMuscularpufdepapadieÎncă nu există evaluări
- Biochima MuschilorDocument7 paginiBiochima MuschilorstikssÎncă nu există evaluări
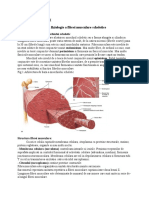
- Fiziologia Muschiului CardiacDocument14 paginiFiziologia Muschiului Cardiacthorscribd100% (7)
- Fiziologia Contractiei MusculareDocument4 paginiFiziologia Contractiei MusculareIoana irimiaÎncă nu există evaluări
- Biofizica Contracţiei MusculareDocument6 paginiBiofizica Contracţiei MuscularenoruletÎncă nu există evaluări
- Morfologia Muschilor Striati 1Document10 paginiMorfologia Muschilor Striati 1AlexColibaÎncă nu există evaluări
- Contractia MuscularaDocument4 paginiContractia MuscularaTudor OlteanÎncă nu există evaluări
- Tesutul MuscularDocument4 paginiTesutul MuscularAriela Săvuţ100% (1)
- M AtpazaDocument8 paginiM AtpazaAlex Grecu100% (1)
- Biofizica Contractiei MusculareDocument11 paginiBiofizica Contractiei MusculareMircea CiobîcăÎncă nu există evaluări
- Bio ChimDocument22 paginiBio ChimEu VictorÎncă nu există evaluări
- FIziologia Aparatului NeuromuscularDocument14 paginiFIziologia Aparatului NeuromuscularLoredana Boghez100% (1)
- Curs - Muschi StriatDocument40 paginiCurs - Muschi StriatDiana Didi Dia100% (1)
- Curs 5 BiofizicaDocument9 paginiCurs 5 BiofizicacurteanancaÎncă nu există evaluări
- Biofizica Contracţiei MusculareDocument4 paginiBiofizica Contracţiei Muscularechamps2009Încă nu există evaluări
- Fiziologia MușchilorDocument12 paginiFiziologia MușchilorVera AndreeaÎncă nu există evaluări
- Structura Si Proprietetile Fiziologice Ale Fibrelor Muschilor StriatiDocument2 paginiStructura Si Proprietetile Fiziologice Ale Fibrelor Muschilor StriatiDaniela IstratiÎncă nu există evaluări
- CURS MUSCHI FIZIO PPTDocument46 paginiCURS MUSCHI FIZIO PPTIrina MureșanÎncă nu există evaluări
- Teorii Si Modele Bio Chi Mice Ale Contractiei MusculareDocument4 paginiTeorii Si Modele Bio Chi Mice Ale Contractiei Muscularesakura.bomboÎncă nu există evaluări
- Curs 5 Muschiul Striat NetedDocument6 paginiCurs 5 Muschiul Striat Netedp3tru_mzqÎncă nu există evaluări
- Fiziologia MuşchilorDocument74 paginiFiziologia MuşchilorXiusa TurcanÎncă nu există evaluări
- Curs 16 AMB IIDocument55 paginiCurs 16 AMB IIDinca Lidia-ElenaÎncă nu există evaluări
- Curs 2 InimaDocument20 paginiCurs 2 InimaAttila SzakacsÎncă nu există evaluări
- Contracția Musculara BF FIMDocument16 paginiContracția Musculara BF FIMVlad ȘtefanÎncă nu există evaluări
- Sistemul NeuromuscDocument67 paginiSistemul NeuromuscAlexandru PurcelÎncă nu există evaluări
- Cursul IV, Note de Curs Neuro-MuscularDocument16 paginiCursul IV, Note de Curs Neuro-MuscularYusef Ceva100% (1)
- Citoplasma CelularaDocument16 paginiCitoplasma CelularaEnea DanielaÎncă nu există evaluări
- Biofizica Aparatului LocomotorDocument10 paginiBiofizica Aparatului LocomotorAurel BoleaÎncă nu există evaluări
- Fiziologie LP NR 3 Anul IDocument13 paginiFiziologie LP NR 3 Anul IAnna MariaÎncă nu există evaluări
- Cursul 4 FizioDocument5 paginiCursul 4 Fiziodanyela0808Încă nu există evaluări
- Curs 5Document14 paginiCurs 5sham harbaÎncă nu există evaluări
- ElectromiogramaDocument6 paginiElectromiogramaAna Maria100% (1)
- 31 Filamentele de ActinaDocument10 pagini31 Filamentele de ActinaAlina Calmuc100% (1)
- 2-Biofizica Contractiei MusculareDocument33 pagini2-Biofizica Contractiei MusculareClaudiu ArdeleanÎncă nu există evaluări
- Sistemul MuscularDocument9 paginiSistemul MuscularDiana AndreeaÎncă nu există evaluări
- Contractia MuscularaDocument12 paginiContractia MuscularaChriss ChristinaÎncă nu există evaluări
- Compoziţia Chimică Generală A Organismului Uman EsteDocument4 paginiCompoziţia Chimică Generală A Organismului Uman EstedenisemichelaÎncă nu există evaluări
- BiocelDocument40 paginiBiocelAdelina MusaÎncă nu există evaluări
- Mecanismul de ContractieDocument1 paginăMecanismul de ContractieDiana ButacuÎncă nu există evaluări
- Curs Neuromuscular-Radu BragaDocument14 paginiCurs Neuromuscular-Radu BragaVicențiu Răzvan VornicuÎncă nu există evaluări
- Structura Tesutului MuscularDocument8 paginiStructura Tesutului MuscularAna-Maria TanasăÎncă nu există evaluări
- Notite AnatomieDocument20 paginiNotite AnatomieStepByStepÎncă nu există evaluări
- Curs Fiziologia Mușchilor Striați Și Netezi-34826Document57 paginiCurs Fiziologia Mușchilor Striați Și Netezi-34826Alina IlovanÎncă nu există evaluări
- Fiziologie TotDocument34 paginiFiziologie TotbamsweÎncă nu există evaluări
- 1.5. Fiziologia Fibrei MusculareDocument82 pagini1.5. Fiziologia Fibrei MusculareAlina Topor Psi-MedicalÎncă nu există evaluări